


1 Dep. de Producción Vegetal, Fac. de Agronomía, Univ. de Buenos Aires, Av. San Martín 4453, Capital Federal (C1417DSE), Argentina. www.agro.uba.ar. E-mail bgambin@agro.uba.ar, otegui@agro.uba.ar.
2 Pioneer Hi-Bred Int., 18285 County Road 96, Woodland, CA 95695-9340, U.S.A. www.pioneer.com. E-mail lucas.borras@pioneer.com.
The curvilinear relation between plant growth rate around silking (PGR) and kernel number in maize implies that, from a given threshold, the intrinsic efficiency of the plant to set kernels declines as PGR increases. This suggests that improved growing conditions around silking increase the assimilate availability per kernel during the first grain-filling phase. So, we hypothesize that increased PGR not only results in higher kernel numbers but also in higher potential kernel weight (KW) at the end of the lag phase. Since maize kernels seem to grow under conditions of sufficient assimilate availability during the effective grain-filling period, this potential KW is usually attained. Thus, we conducted an experiment with twelve genotypes differing in final KW to evaluate if PGR around silking could help explain differences in final KW within and between hybrids. We estimated potential KW from kernel water content values at the end of the lag phase. Each genotype showed a different response pattern in potential KW to changes in PGRs. However, we validated our hypothesis stating that within each genotype, higher PGRs were related to higher potential KWs because of an increase in plant growth rate per kernel during the kernel set period.
Key words
Zea mays L., maize, plant growth rate per kernel, kernel weight, kernel water content, genotypic differences.
Introduction
In major extensive crops yield is mainly determined by the number of harvested seeds per unit land area, while individual seed dry weight plays a secondary role in most cases (Fisher, 1975). The seed set period, generally around flowering, is usually defined as a stage during the crop growing cycle when yield is mostly source limited. Once sink size is defined, a source-sink co-limitation prevails, and the relative importance of this co-limitation varies depending on the species and environmental conditions during grain filling (Borrás et al., 2004). It has been shown that maize kernels normally grow under saturated conditions of assimilate availability along the effective grain-filling period, and it is only during the first stages of grain filling that they may experience source limitations under normal growing conditions (Borrás et al., 2004). This led us to speculate that at the end of the first grain-filling phase, known as the lag phase, maize plants have already set not only a number of kernels (Andrade et al. 1999) but also a potential weight for that particular growing condition. Most maize crops are able to realize this potential kernel weight (KW) at the end of the cycle if no source restrictions are imposed from the lag phase onwards.
The typical curvilinear response of kernel number per plant to plant growth rate around silking (PGR) found in maize (Andrade et al., 1999) implies that the intrinsic efficiency of the plant to set kernels declines as PGR increases, especially at PGRs larger than 1 g plant-1 day-1 (Vega et al., 2001). This means that PGR per kernel increases as plant growth increases during this period. Understanding this, we currently hypothesize that high PGRs around flowering not only determine high kernel numbers but also high KWs. Examples of this are (i) stand density treatments, where plants at the low stand densities establish higher kernel numbers and higher KWs than their counterparts at high plant populations (Borrás et al., 2003), or (ii) drought treatments only around the flowering period, where drought reduces kernel number as well as final KW (Chapman and Edmeades, 1999). Another clue that makes us speculate that the PGR per kernel during the seed set period is behind differences in final KW are the results from pollination treatments performed at this stage and aimed to alter only the number of kernels set by the plant (Borrás and Otegui, 2001). For the same plant growing condition (i.e., plant growth rate), KW decreased whenever kernel number per plant was increased, and increased whenever the number of kernels was reduced. Therefore, taking into account the curvilinear relationship between kernel number per plant and PGR, we hypothesize that the potential KW increases under improved growing conditions, mainly because more source is available for each kernel during the lag phase.
So far, hypotheses dealing with potential KW have been very difficult to address because of the intrinsic variability of the classic kernel sink potential estimators (like the number of endosperm cells or of starch granules) and the difficulty for measuring them. Recently, a new methodology for estimating potential KW in a fairly easy and accurate way has been developed using kernel water relations (Borrás and Westgate, 2002). It is based on the fact that final KW is related to the maximum amount of water kernels achieve at mid grain filling (Borrás et al., 2003), and that not only KW but also the development of water content can be modeled with kernel moisture content. So, using moisture content as a benchmark for the estimation of the developmental status of kernels at any moment along the grain-filling stage, it is possible to predict potential KW early in kernel development. This methodology has been validated for a very wide range of environments and genotypes (Borrás and Westgate, 2002).
Objectives
- To study if higher PGRs around silking are related to higher potential KWs due to a higher plant growth rate per kernel.
- To evaluate if genotypic differences in final KW are related to differences in the potential kernel weight established at the end of the lag phase.
Materials and Methods
Twelve Argentine commercial genotypes (Ax842 MG, Ax610 MG, Ax820 MG, Ax878 MG, Xa0675 MG, Ax882 MG, Ax888 CL-MG, Ax800 MG, Ax832 CL-MG, Ax889 MG, Ax890 MG, Ax877 MG) from Nidera Argentina, of contrasting final KW and endosperm type, were used. Eight plants within each of the three replicates were tagged 15 days before silking, and their biomass increase around silking was predicted using non-destructive allometric models (Vega et al., 2001). Based on the methodology from Borrás and Westgate (2002), kernel water content on 10 to 20 days after silking (around 83% moisture content) was used for the prediction of maximum kernel water content. This was performed by the application of the parabolic equation of percent maximum water content and moisture content to data obtained from fifteen kernels. Potential KW was computed from the linear relationship between maximum water content and final KW (Borrás et al., 2003).
Results and Discussion
Each genotype explored a wide range of PGRs around silking (from 2 to 5 g plant-1 day-1), and showed a different response pattern in potential KW to changes in PGRs. This relationship is represented in Figure 1A for only two genotypes. In both cases, PGR increases during the flowering period resulted in an increase in the potential KW achieved at the end of the lag phase, as PGR per kernel augmented with increasing PGRs (Figure 1B). When these two genotypes were compared, however, a single response pattern of potential KW to PGR per kernel did not fit the whole data set, as similar abscissa values yielded contrasting potential kernel weights (Fig. 1A and B).
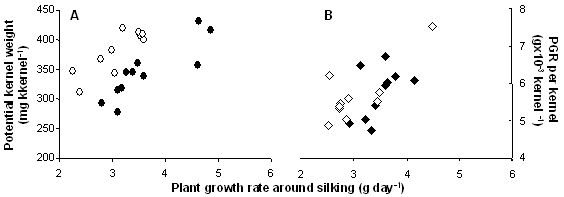
Figure 1. Relationship between potential kernel weight (A) and plant growth rate per kernel (B) with plant growth rate around silking for hybrids Ax 878 MG (open symbols) and Ax 877 MG (closed symbols).
When data from all genotypes were pulled together, differences in potential KW (from 288 to 360 mg kernel-1) at 15 days after silking were evident (Figure 2). These values, however, overestimated KW records actually registered at the end of grain filling for each genotype (Figure 1). Two possible reasons for this trend are: (i) that some genotypes experienced a source limitation during the effective grain-filling period, which hindered the achievement of the potential kernel weight established early in development, or (ii) an error in the estimation of potential kernel weight by using the model described by Borrás and Westgate (2002). We believe the latter is the main reason, as evidenced in Figure 3. Between 80 and 88% moisture content (end of the lag phase), water content values in our study were higher than the adjusted parabolic equation obtained by Borrás and Westgate (2002), and may have overestimated our potential kernel weight values (Figure 2). This fact shows the need of verifying the genotypic range for which the model was built as a source of error for future uses. Also, although there might have been an overestimation in the potential KW calculation, this overestimation was the same for all genotypes, as all of them were clearly showing the same pattern of response, especially in the range at which potential kernel weight is estimated (80-88% moisture content).
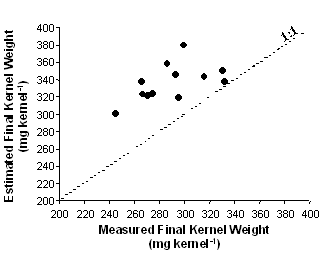
Figure 2. Measured and estimated final kernel weight from twelve genotypes differing in kernel weight and endosperm type.
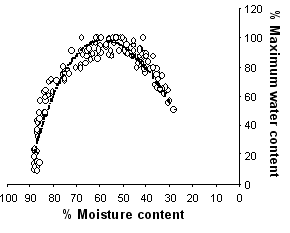
Figure 3. Percent of maximum water content and moisture content from the twelve genotypes tested. The dotted line shows the equation fitted in Borrás and Westgate (2002).
Conclusions
Results shown in Figure 1 validated our hypothesis stating that, within each genotype, higher PGRs were related to higher potential KWs at the end of the lag phase because of an increase in plant growth rate per kernel. For the first time, our study demonstrated that potential kernel weight at the end of the lag phase and plant growth rate around silking are related. A larger number of environments and genotypes are needed to state more robust conclusions, but so far present results seem to support our working hypothesis.
Caution has to be taken when extrapolating the model to calculate potential kernel weight from water content early in development from one set of genotypes to another one, as it seems that not all genotypes have the same water developmental pattern.
References
Andrade FH, Vega C, Uhart S, Cirilo A, Cantarero M and Valentinuz O (1999). Kernel number determination in maize. Crop Science 39, 453-459.
Borrás L and Otegui ME (2001). Maize kernel weight response to postflowering source-sink ratio. Crop Science 49, 1816-1822.
Borrás L and Westgate ME (2002). Relations between dry matter and water content in maize kernels: implications in the control of final kernel weight. XI Latin American Plant Physiology Meeting, October 22-25, Punta del Este, Uruguay, pg. 56.
Borrás L, Westgate ME and Otegui ME (2003). Control of kernel weight and kernel water relations by post-flowering source-sink ratio in maize. Annals of Botany 91, 857-867.
Borrás L, Slafer GA and Otegui ME (2004). Seed dry weight response to source-sink ratio manipulations in wheat, maize and soybean: a quantitative reappraisal. Field Crops Research 86, 131-146.
Chapman SC and Edmeades GO (1999). Selection improves drought tolerance in tropical maize populations. II. Direct and correlated responses among secondary traits. Crop Science 39, 1315-1324.
Fisher RA (1975). Yield potential of dwarf spring wheat and the effect of shading. Crop Science 15, 607-613.
Saini HS and Westgate ME (2000). Reproductive development in grain crops during drought. Advances in Agronomy 68, 59-96.
Vega CRC, Andrade FH and Sadras VO (2001). Reproductive partitioning and seed set efficiency in soybean, sunflower and maize. Field Crops Research 72:163-175.

Search Site
- Crop Science
- Help/Feedback
- Contact us
- Sitemap