


1New Zealand Institute for Crop & Food Research, Private Bag 4704, Christchurch, NZ www.crop.cri.nzEmail wilsond@crop.cri.nz
2New Zealand Institute for Crop & Food Research, P.O. Box 85, Hastings, NZ
Diverse types of forage brassica crops are sown throughout the year in a wide range of climates and soils to supplement pastures in grazing animal production systems. A potential yield model is being developed for these crops as the basis of a forecasting system to optimise crop management. It is similar in principle to existing models for other crops. Canopy development is simulated by calculating the leaf area of an average plant each day, and then accumulating over all plants in the population. Leaf area calculations account for the production and loss of successive leaves. Daily biomass production is the product of the amount of radiation intercepted by the canopy and a constant radiation use efficiency. New biomass is distributed to leaf, stem and root fractions using empirical partitioning coefficients. The model accurately predicts canopy development, biomass production, and biomass distribution for two different brassica types (Pasja and kale, which produce mainly leaf and stem biomass respectively). Further model development is needed to cope with variable plant size which leads to self-thinning in brassica types such as kale which have relatively low populations, large average plant size and slow development rate. More research is needed to check the wider applicability of the model, to define the distinguishing features of other brassica types, and then to adapt the model for them.
A yield model is being developed for a computer-based forecasting system to optimise management of brassica crops grown to supplement pastures in grazing animal production systems.
Keywords
yield prediction, canopy development, radiation use efficiency, biomass production, partitioning
Introduction
Forage brassicas are grown widely to supplement pastures in grazing animal production systems. In New Zealand, they comprise the largest area of cultivated crops, with about 250,000 ha grown annually. The forage brassicas have diverse growth types and encompass various species and inter-specific hybrids. They include crops that produce primarily root biomass (e.g. swede (Brassica napus L.) and turnip (B. rapa L.)), others that produce mainly leaf and stem (e.g. kale (B. oleracea L.) and rape (B. napus L.)), and some that produce much leaf but little stem or root (e.g. Pasja, a fast-growing, early-maturing turnip-Chinese cabbage hybrid).
Besides their varied types, forage brassica crops are sown throughout the year and are grown in a wide range of climates and soils. This diversity makes it difficult to specify optimum management. For example, it is difficult to predict fertiliser requirements because yield responses to fertiliser application, which depend on potential yield and soil fertility, are very variable. We are developing a forecasting system that will use a new approach to predict these responses for forage brassica crops (Reid 1999; Reid 2002; Reid et al. 2002). It comprises two elements: (a) a model that predicts potential yield, with no nutrient or water limitations, which is driven by radiation and temperature and also accounts for the effects of sowing date and plant population and (b) a model that reduces yield below the potential according to the availability of each macro-nutrient from both soil and fertiliser sources.
No potential yield models exist for forage brassicas. Therefore, as a first step towards the forecasting system, in this paper we describe a new generic radiation and temperature driven model for the diverse range of forage brassica crops. We also present results from field experiments that were conducted to produce information for developing the model, and for testing it by comparing predictions with results from independent experiments.
Methods
Initial model development has focussed on two brassica types, Pasja and kale, with contrasting biomass distribution characteristics. Field experiments were conducted with both crops to quantify key features of the three main elements of the model: canopy development and radiation interception, biomass production, and biomass partitioning. Each crop was sown at two locations on three dates spanning the usual planting period, and at three plant populations which encompassed the usual range. The crops were managed intensively, using best practices for forage brassicas, to minimise biotic and abiotic stresses and thereby to ensure maximum expression of yield potential. Data from experiments at one location were used for model development, and results from experiments at the other location were used for testing the model.
Numbers of fully expanded and senesced leaves on tagged plants were counted regularly, and area profiles were determined by measuring the area per leaf starting from the bottom of representative plants in each population treatment. Biomass production and partitioning were measured by harvesting quadrats at regular intervals and measuring the dry weights of leaf, stem and storage root components. Measurements and subsequent analyses of results were adapted for Pasja and kale to account for their contrasting features. One difference was that Pasja consisted mainly of leaves and small storage roots, while kale comprised leaves and a large storage stem component. Another difference was that Pasja had uniform populations of plants which ranged from about 60 to 180 per m2 and rapidly produced full leaf canopies. In contrast, kale had populations of more diverse plants which ranged from about 40 to 100 per m2, produced leaves at a slower rate, and took longer to develop full canopies. As a result, the growth and development of individual kale plants had much more influence on crop performance per unit area than individual Pasja plants.
Model description
The model is similar to potential yield models for other crops (e.g. Jamieson et al. 1998; Muchow et al. 1990), but without the partitioning to seeds that is associated with reproductive development because brassicas are biennials. They are grown for forage in the first year, and develop reproductively in the second year after flowering has been initiated during the winter. The model describes only the vegetative production year.
Canopy development is simulated by calculating the leaf area of an average plant each day, and then accumulating the total leaf area for all plants in the population. Leaf area on the average plant is calculated by accounting for the production and loss by senescence of successive leaves. The procedure is more complex for kale because of the heterogeneity in the plant population and its slower rate of leaf area development. The heterogeneity occurs because plants germinate and emerge over a substantial period. The resulting differences among plants persist, and they are accounted for by dividing the population into cohorts. Each cohort, which consists of plants with similar leaf number, leaf area and biomass, is simulated separately. The cohort model is described in a companion paper (Zyskowski et al. 2004). The rules for canopy development are:
- There is no limit on the number of leaves that a plant can produce in the first year.
- The rate of leaf appearance responds linearly to temperature above a 0°C base. Leaf appearance rate is the principal distinguishing feature of canopy development between the two crop types:
- Pasja produces leaves, and therefore develops a full canopy, much faster than kale. The mean leaf appearance rate for Pasja was 0.016 leaf per °C day (i.e. phyllochron about 62 °C days) (Figure 1). There was evidence that leaves were produced faster at lower populations, perhaps because plants were under less competitive pressure for resources.
- Kale produces leaves at just over half the rate of Pasja, with a value of 0.009 leaf per °C day (i.e. phyllochron about 110 °C days) (Figure 1).
- area per leaf depends on leaf position up the plant, population and (in kale) size cohort. For example, in both crops, at a normal plant population the full leaf size increases from leaf 1 to about leaf 12 and, thereafter, size is constant. Competition for resources starts earlier, so the area of comparable leaves is smaller, as population increases. In kale maximum leaf size ranges from 0.05 m2 for large-cohort plants at low population to 0.01 m2 for small-cohort plants at high population.
- Expansion of each leaf to its full size is defined by a dimensionless “shape” factor which is 0.004 for kale and 0.002 for Pasja (Figure 2).
- leaf has a non-linear, population-dependent thermal duration from first appearance until it starts to senesce. At normal populations of 120 and 70 plants per m2, the thermal duration of each leaf is 258 and 285 °C days for Pasja and kale respectively. It then senesces at a constant rate in thermal time (0.002 m2 per °C day) (Figure 2).
When these rules are accumulated, each plant has a predictable number of leaves and leaf sizes at any time and population. Total area is calculated by acumulating the areas of all leaves on all plants in a population.
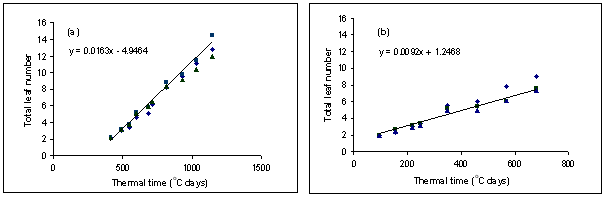
Figure 1. Leaf appearance in thermal time for (a) Pasja and (b) kale. Data are from three sowing dates and the fitted line is for a medium plant population.
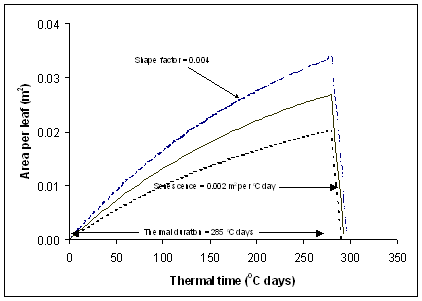
Figure 2. Expansion of individual leaves in thermal time from emergence for kale. The pattern of expansion is defined by the “shape” factor and the lines illustrate the effects of different leaf position or plant population.
Daily biomass production (g/m2) is calculated as the product of the amount of radiation intercepted by the canopy (MJ/m2) and a constant radiation use efficiency (1.2 g/MJ). Radiation intercepted is calculated as a function of canopy leaf area using Beer’s Law with an extinction coefficient (k) of 0.75. Cumulative radiation intercepted is not very sensitive to the value of k because, except for kale at low populations, the leaf canopies of both crops develop quickly to full interceptance.
Besides contrasting canopy development, different biomass distribution is the main distinction among the brassica types. In the model, constant proportions of new biomass are distributed daily to leaf, stem and root components using empirical partitioning coefficients. The coefficients for Pasja are 95% to leaf (20% lamina and 75% petiole) and 5% to root. Leaf, stem and root coefficients for kale are 35%, 60% and 5%. Biomass released from senesced leaves is redistributed to storage in roots and stems in Pasja and kale respectively.
Results
The model accurately predicted leaf area index at three plant populations in the independent experiments for both Pasja and kale (Figure 3). There was also good agreement between simulated and measured values of total biomass production and partitioning of the biomass to leaf and root for Pasja and to leaf and stem for kale (Figure 4). In the case of Pasja, which is usually multi-grazed at about 60-day intervals, the model also accurately predicted leaf and biomass production following a simulated defoliation.
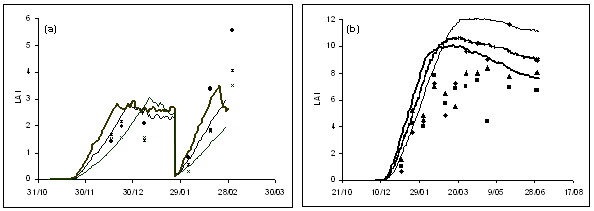
Figure 3. Simulated and measured leaf area index (LAI) for (a) Pasja and (b) kale. Simulations and data for each crop are from one sowing date and three plant populations and, for Pasja, include regrowth following a simulated grazing.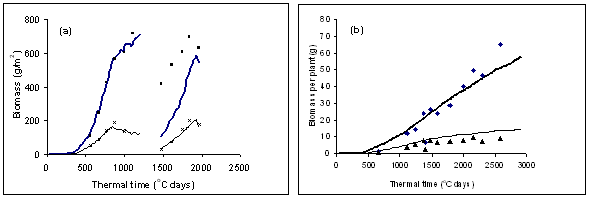
Figure 4. Simulated and measured total (top) and leaf lamina (bottom) production for (a) Pasja and (b) kale. Simulations and data for each crop are from one sowing date and a medium plant population and, for Pasja, include regrowth following a simulated grazing.
Conclusion
This model provides a framework for predicting potential yield of forage brassicas. Preliminary testing shows that it accurately predicts potential production by Pasja and kale crops grown at a range of plant populations. These crops are sown at various times of the year in a wide range of climates and soils, and more testing is needed to check the wider applicability of the model. Different brassica types are distinguished mainly by contrasting canopy development characteristics and different distribution of biomass. More research is needed to define these features for other types, and to adapt the model for them. Further work is in progress to cope with variable plant size which leads to self-thinning in brassica types such as kale which have relatively low populations, large average plant size and slow development rate (Zyskowski et al. 2004).
References
Jamieson PD, Semenov MA, Brooking IR and Francis GS (1998). Sirius: a mechanistic model of wheat response to environmental variation. European Journal of Agronomy 8, 161-179.
Muchow RC, Sinclair TR and Bennett JM (1990). Temperature and solar radiation effects on potential maize yield across locations. Agronomy Journal 82, 338-343.
Reid JB (1999). Forecasting nutrient responses in annual crops. Agronomy New Zealand 29, 69-72.
Reid JB (2002). Yield response to nutrient supply across a wide range of conditions 1. Model derivation. Field Crops Research 77, 161-171.
Reid JB, Stone PJ, Pearson AJ and Wilson DR (2002). Yield response to nutrient supply across a wide range of conditions 2. Analysis of maize yields. Field Crops Research 77, 173-189.
Zyskowski RF, Wilson DR and Maley S. (2004). A cohort model for simulating forage brassica crops with variable plant size. This Volume.

Search Site
- Crop Science
- Help/Feedback
- Contact us
- Sitemap