


Graduate School of Agriculture, Kyoto University, Kyoto 606-8502, Japan, http://www.kyoto-u.ac.jp. email: horiet@adm.kais.kyoto-u.ac.jp
The annual rate of rice yield increase in the world declined from 2.7 % in the 1980s to 1.1 % in the1990s. The continued world population increase requires resumption of the previous rate. The objectives of this paper are to review and assess rice production technologies for increased yield in the past and the current challenge on the basis of crop physiology and agronomy, and to discuss the way to increase irrigated rice yield to fulfill the expanding demand. Field experiments conducted in Kyoto, Japan and Yunnan, China showed that the best recent Chinese hybrid has a yield potential about 10 % higher than the best recent inbred cultivar in Japan. This result and a review on recent challenges in breeding suggested only moderate increases in the yield potential of rice genotypes in the coming decades. Contrary to the general understanding, the rapid rice yield increase in central Japan from the mid 1950s to mid 1970s was achieved mostly by improved crop and resource management. This and the fact that farmers’ contest-winning yields during 1950s and 1960s nearly doubled the current average in Japan imply that current crop and resource management is exploiting only part of large yield potential of rice.
Some of the ways to increase yields may include components of the system of rice intensification (SRI). The extremely high yields in SRI are incredible but its elements, which have been studied and practiced in Japan for the past 50 years, may lead to yield increases. The practice of transplanting one or two young seedlings per hill has advantages in reducing transplanting injury and increasing tiller and root numbers on lower nodes. Such advantages can be realized under direct-seeding systems, where they are applicable. The practice of applying a large amount of compost and intermittent irrigation were also adopted by many of the contest-winning farmers in the 1950s and 1960s. These practices increase roots in deeper soil layers, maintain their activities and presumably promote nitrogen (N) uptake at later stages. Remarkable progress has been made in improved N management; agronomic efficiency of N increased from about 15 kg kg-1 for a single dose at transplanting to 40 kg kg-1 for banded controlled release fertilizer. All of these technological elements will contribute to increased yield when they are rationally integrated into a system that is adaptable to regional environment.
Genetic improvement triggered most of the previous yield increase of tropical rice but relatively little of the yield increases in Japan, where improved crop management provided most of the gains. Future genetic improvements in global rice yield are likely to be modest but there are several new crop management practices that can significantly increase yield.
Key Words
Hybrids, aerobic culture, SRI, controlled release fertilizers, early transplanting, plant density
Introduction
World rice production nearly doubled in the two decades from the mid 1960s to the mid 1980s. Approximately 80% of the production increase was achieved by higher yield, through gradual replacement of traditional cultivars by modern cultivars developed in rice research centers, supported by public investment for expansions of irrigation infrastructure and expansion of credit facilities (Hossain, 1999). The dramatic yield increase followed the technological innovations referred to as the Green Revolution. However, by the mid 1980s, Green Revolution production technologies had expanded throughout Asia, and rice yield increase slackened, as reflected by the sharp drop of the annual yield increase rate from 2.7% in the 1980s to 1.1% in the 1990s. Despite this, continued expansion of world population and economic development will require 56% more rice production in the next three decades (International Rice Research Institute: IRRI, 1997). This, together with fertile paddy fields being converted for more profitable crops and urban infrastructure in many countries, requires resumption of rice yield increase to a comparable level to that in 1980s. This needs to be achieved through efficient use of water, nutrients and other resources under the current situations of scarce resources and increasing demand for conservation of the natural environment.
The challenges to crop scientists are to increase yield and resources use efficiency both in irrigated and rainfed lowland rice cultures, by breeding high-yielding genotypes and developing more efficient crop management technologies. Breeding challenges are to create new plant type (NPTs, Khush and Peng, 1996), F1 hybrids (Yuan, 1994; 2001) that break through the current yield limit of irrigated rice, and superior genotypes that increase yield under resource-poor and unstable rainfed conditions (Fukai and Cooper, 1995; Pantuwan et al., 2002). Challenges of crop and resource management, are the system of rice intensification (SRI, Uphoff and Randriamiharisoa, 2002; Stoop et al., 2002), improved water managements of intermittent irrigation (Shi et al, 2002; Belder et al., 2002), aerobic rice culture (Wang et al., 2002; Lin et al., 2002), and the use of controlled release fertilizers (Shoji and Kanno, 1994).
Assessment of those diverse challenges, in view of their potential for contribution to rice yield increase, have not fully been made. This review assesses current achievements and , on the basis of crop physiology and agronomy, the potential and feasibility of these technologies to increase yield of irrigated rice, and points out strategy that will resume yield increases to a comparable rate to that in the 1980s. This paper first reviews historical rice yield increase and the associated factors, then assesses the current yield potential increase of genotypes and high-yielding rice culture technologies, and discusses the strategy toward increased rice yield.
Historical Courses of Rice Yield Increase in Selected Countries and Associated Factors
Time courses of rice yield increase in selected countries
To clarify whether rice yield has plateaued over Asian countries and at what level, we examined historical time courses of the yield for China, Japan, Indonesia and India. (Fig. 1, from FAO, 2004). Fig. 1 also represents, for comparison, the record yield of Japanese farmers’ rice contest for the period 1950 to 1963 during which the contest was made. The periods of rapid yield increase were different among the countries examined: mid 1950s to mid 1970s in Japan, late 1960s to mid 1980s in Indonesia and mid 1970s to mid 1990s in China and India. These periods may be called Green Revolution periods of the respective countries. The technologies that enabled the Green Revolution are generally accepted to be adoption of modern high yielding cultivars and irrigation, use of N fertilizers and chemicals to control weeds, pests and diseases ( e.g. Hossain, 1999). In China, diffusion of hybrids contributed for the yield increase, because hybrids’ yield exceeded inbred cultivars by 15 % (Yuan, 2001) and diffusion of hybrids reached to nearly 50% in 1990s. However, Japanese yield increase was somewhat different from the general understanding, because almost all lowland rice was already irrigated in 1950s and the contribution of cultivars was relatively small. The technologies associated with Japanese rice yield increase is analyzed in the subsequent section.
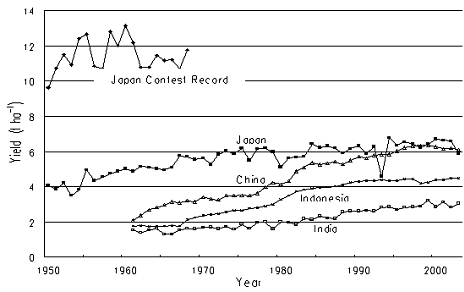
Fig. 1. Changes with year in rice yield of selected Asian countries and in yield contests in Japan from 1950 to 1963.
It is commonly recognized over the four countries examined that their yield increases have slackened since the termination of the respective Green Revolution periods. However, the near-plateau yields differ among the four countries: about 6.5, 4.5 and 3.0 t ha-1 for China and Japan, Indonesia, and India, respectively. This yield difference among the countries reflects percentage of area irrigated. Rainfed rice that occupies nearly 50% of Asian rice land was left outside the Green Revolution and its yield remains as low as about 2t ha-1 in most regions.
Another important point in Fig. 1 is that the contest-winning yield in Japan had reached 12t ha-1 in 1950s, which was almost triple the national average yield at that time and nearly double the current yield in Japan. The major technologies employed by many of the contest-winning farmers were drainage, deep plowing, application of a large amount of manure, and water management systems of midseason drainage and/or intermittent irrigation (Fuke, 1961; Kawata, 1976). The effects of these technologies on rice growth and yield are the subject for the analysis in the following section.
Factors associated with the yield increase in Japan since 1950s
As shown in Fig. 1, Japanese rice yield increased by about 50% from the mid 1950s to mid 1970s, and thereafter the rate of increase slackened. The motive force for the rapid yield increase was strong demand for rice self-sufficiency because Japan faced severe food shortage after the Second World War. During this period intensive research and farmers’ trials, including the rice yield contests, formed the basis of the present rice production technologies in Japan. Analysis of factors and technologies that enabled the yield increase provide indications for future rice yield increase in Asia. Hasegawa et al. (1991) and Hasegawa and Horie (1995) analyzed the effects of cultivars and agronomic technologies on yield increase from 1960 to 1980 in the Kinki district in Japan. They suggested that the contribution of cultivar improvement for the rapid yield increase during this period was relatively small. We here extend this analysis for the period from 1950 to the present for Shiga prefecture in central Japan. Many new cultivars were bred and introduced to Shiga in this period. To separately evaluate the cultivar effect on the yield increase from other technological effects, we employed the method of Feyerham et al. (1984), as Hasegawa et al. (1991) did. Field tests for evaluating rice cultivars have been conducted in many agricultural stations in Japan including Shiga. We utilized those data from the Shiga Pref. Agr. Exp. Stations for the period from 1950 afterward. Using those data, the difference in yielding ability between a given cultivar and a standard cultivar is presented as the differential yielding ability (DYA) which is calculated by:
DYA ί= 1/ n ![]() (Y(c)ijk – Y(s)ijk) (1)
(Y(c)ijk – Y(s)ijk) (1)
where n is the total number of data pairs for comparison between cultivar, C, and the standard, S, Y(c)ijk and Y(s)ijk are yields in ith treatments at the jth location in kth year for the tested cultivar, C, and the standard, S, respectively. Cultivar Nipponbare was adopted as the standard cultivar, because it was represented in almost every performance test analysed since 1963 when it was released. For very old cultivars, with which cv. Nipponbare was rarely tested, cv. Kinmaze was used as the secondary standard and the temporary DYA’s for the old cultivars against cv. Kinmaze were translated to that against cv. Nipponbare through DYA of Kinmaze against Nipponbare.
Many cultivars were simultaneously grown in a given region and year, and the constituent cultivars changed year by year. To evaluate the cultivar effect on regional yield increase under these circumstances, regional differential yielding ability (RDYA) for a given year was calculated by,
RDYA= ![]() ai DYAi /
ai DYAi / ![]() ai (2)
ai (2)
where ai indicates the percentage planted area of the ith cultivar, and ![]() the total number of cultivars grown in the region in a given year.
the total number of cultivars grown in the region in a given year.

Fig. 2. Differential yielding ability (DYA) of 34 widely-grown cultivars in Shiga Prefecture in the central part of Japan during the past 50 years plotted against year of release. The star indicates the standard cultivar Nipponbare.

Fig. 3. Increase in the average rice yield (Yield) since 1950 for Shiga Prefecture compared with the change in regional DYA (R-DYA) (A) and with the average planting date (B), and changes in the number of power tillers and rate of N application (C). Yield (7 year moving average) and planting date are from the Crop Production Year Book of Japan. R-DYA is weighted mean of DYA of cultivars based on the planted ratio of each cultivar in this region and presented by the difference from the value at 1950.
Fig. 2 shows changes in cultivar DYA as a function of its release year in Shiga. An increase in DYA is recognized before 1960s, indicating that yields of newly released cultivars exceeded that of older cultivars during this period. From around 1960, however, cultivar DYA plateaued, indicating no substantial yield increase for cultivars released after this year. This implies that the breeding target changed at that time from one yield to multiple targets that included quality and pest and disease resistances, as well as yield.
Reflecting the time course of cultivar DYA, RDYA increased only until the early 1970s and thereafter it plateaued and then decreased (Fig. 3). Although the increase of RDYA slowed during the 1960s, the actual yield in Shiga kept increasing by 1.5% per year up to around 1975. When the changes in RDYA and the actual yield were compared, the cultivar effects on the yield increase in Shiga prefecture during the 1950 to 1975 period was estimated to be only 40% and that for the later half of the period was as low as 20%. Thus a major part of the yield increase in this prefecture has to be ascribed to factors other than cultivars. Increase in the percentage irrigated area as in other Asian countries was not the case in Shiga, because almost all paddy fields were already irrigated in 1950. In this period, especially during the later half, the average planting date became progressively earlier from mid June to early May (Fig. 3b) and this change coincided closely with the yield trend. It is evident that the earlier planting favored yield formation, presumably through the prolonged growth period. One technological factor that supported the early planting was the protected nursery. This technique to protect young seedling from low temperature was established in 1942 and fast spread in the whole country since the mid 1950s (Hayashi, 1961). The second factor was mechanization of land preparation. The ratio of farms equipped with power tillers in Shiga was only 5% at 1958 and it grew consistently to near 100% in the late 1970s. This change greatly improved efficiency of preparation of paddy fields and enabled the earlier planting of rice.
The rate of N fertilization also increased from about 60kg ha-1 in the mid 1950s to about 100 kg ha-1 around 1970. This also contributed to the yield increase in the period before 1975, as well as the early planting culture. From mid 1970s, the yield increase in Shiga has slackened, reflecting rice overproduction in Japan. Since around 1970, rice production in Japan has been directed to better quality and higher efficiencies in labor productivity and resource use. In particular, N fertilization rate began to decrease in 1980, after which the yield level was maintained. This is a consequence of improved fertilization methods and will be discussed later.
The analysis on the yield increase in Shiga clarified that the sharp yield increase in the period from 1950 to 1975 was achieved mainly by improved crop and resource management. Also, it is clear that the reason that contest-winning yield in the 1950s and 1960s were nearly double the average yields was solely due to better management practices. These facts suggest that there is still a large room for increasing rice yield by better management of the crop and resources.
Yield Potential Increase of Rice Genotypes
The yield potential of recent genotypes
Crop yield potential is the maximum yield of a crop genotype growing under no biotic and abiotic stresses and with an optimum supply of resources. The greatest improvement in yield potential was due to the release of short-stature japonica genotypes with the semi-dwarf gene from cv. Ginbouzu, which was selected in Japan in 1907, and indica genotypes, starting with IR8 at IRRI in 1966. These high-yielding genotypes together with expansion of irrigation area and high inputs of fertilizers and agro-chemicals triggered the Green Revolution in rice production in Asia.
In the tropics, however, rice yield potential has not increased substantially in the past three decades since the Green Revolution began and the gap between yield potential and farmers’ yield has narrowed (Peng et al., 1999). Efforts have been made to breed genotypes that increase rice yield potential, including the development of new plant types at IRRI (NPTs; Khush and Peng, 1996), super hybrid rice in China (Yuan, 2001) and super-high yielding rice in Japan.
The intended plant characteristics for NPTs were low-tillering capacity with no unproductive tillers, large panicle size, medium plant height, sturdy stems, thick erect leaves with high nitrogen content, 110-130 days growth duration and, as a whole, 30-50% higher yield potential than the existing semidwarf cultivars in tropical conditions (Peng et al., 1994). By introducing genes from tropical japonica to indica cultivars, genotypes with almost all these characteristics were bred. However, the yield potentials of NPTs were not significantly higher than the exciting cultivars, mainly due to poor grain filling (Khush and Peng, 1996; Ying et al., 1998; Horie, 2001). The Japanese Ministry of Agriculture, Forestry and Fisheries (MAFF) initiated super-high yielding rice project in 1981 and released Takanari, an indica-japonica crossbred, in 1990. Breeding of high-yielding hybrid rice, initiated in China in 1970s, have actively been made in many Asian institutions (Virmani, 2001).
Table 1. Above-ground biomass dry weight and yield (14% moisture) of high yielding genotypes grown in Kyoto, Japan and Yunnan, China.

To examine the yield potential of those recent high-yielding cultivars and to identify the traits associated with high-yielding rice, we conducted field experiments comparing cultivars at Kyoto, Japan and Yunnan China. While the Kyoto (35o latitude and 20m elevation) had a typical warm temperate climate, the Yunnan site (26o latitude and 1100m elevation) had a large day-night temperature difference with moderately high solar radiation which produces very high yields (Ying et al., 1998). Details of this experiment were described by Horie et al. (2003b). Table 1 shows the yield data for selected cultivars at Kyoto and Yunnan. The yields of recently bred high-yielding genotypes exceeded those of the check cultivar Nipponbare by different amounts, depending on locations and years. The recently bred Chinese hybrid Liangyoupeijiu (Zou et al., 2003) outyielded Nipponbare by 55%, and the best Japanese inbred cultivar Takanari outyielded it by 38% on average over the locations and years. This indicates that the recent Chinese high-yielding hybrid had a yield advantage of 12% over the best inbred cultivar in Japan. This value is close to the reported yield advantage of about 15% over the best inbred cultivars in China (Yuan, 1994) and 9% at IRRI (Peng et al., 1999). These figures indicate that the recent increase in rice yield potential was not so dramatic as the Green Revolution, but proceeds at a moderate rate.
Yield determination processes and associated traits; perspective of yield potential increase
Horie (2001) reported that difference in yield potential of genotypes and also the difference in yields of a genotype under different environments are reflected in crop growth rate (CGR) during a 2-week period preceding full heading. He pointed out that the CGR during this period determines yield through effects on spikelet number per unit area, single grain mass and potential grain filling (Fig.4). Actual spikelet number per unit area is the difference between number of spikelets that are differentiated and that degenerated. Rice generally differentiates excess spikelets, depending on previous nitrogen (N) uptake (Wada, 1969; Akita, 1989; Hasegawa et al., 1994; Horie et al., 1997).The spikelets then degenerate during the 2-week period preceding full heading, depending on the availability of carbohydrates (Matsushima, 1957; Wada, 1969). Thus, actual number of spikelet per unit area is substantially determined by the availability of carbohydrates during this period. Rice single-grain weight is limited by husk size, which is also determined by the availability of assimilates during this period (Matsushima, 1957; Seo and Ota, 1983).
The 2-week period preceding full heading in rice is the stage when vegetative organs approach maximum growth and nonstructural carbohydrates (NSC) accumulate in culms and leaf sheaths. Generally, the contribution of NSC stored before heading for final grain yield is about 30% (Cock and Yoshida, 1972; Akita, 1989). However, much evidence has been accumulated to indicate that NSC accumulated before the onset of grain-filling plays a more important role in rice grain-filling than merely compensation for a shortage of assimilates during the grain-filling (Takeda et al., 1980; Seo and Ota, 1983; Akita, 1989; Nakamura et al., 1992; Sumi et al., 1996; Tsukaguchi et al., 1996; Horie et al., 1997). It is reported for rice (Nakamura et al., 1992) and wheat (Radley, 1978; Signh and Jenner, 1984; Feught and Hofner, 1985) that final grain size is closely related to endosperm cell number, which in turn is controlled by the supply of substrate during its formation. For rice the final number of endosperm cells is clearly determined within about the initial 10 days after flowering (Hoshikawa, 1967). Photosynthetic production during the initial 10 day of grain-filling is usually not enough to supply the necessary amount of carbohydrate for all the spikelets in a panicle for their full development in endosperm cell number, and it is more so in rice with a large number of spikelets. These previous studies suggest that higher CGR during the 2-week period preceding full heading enables more accumulation of NSC, which in turn contributes to a higher grain-filling percentage by determining endosperm cell number.

Fig. 4. Schematic presentation of the processes through which crop growth rate (CGR) during the latter half of the reproductive period (14 to 0 d before heading, DBH) has critical effects on yield (Horie, 2001). NSC = nonstructural carbohydrates.
There are indications that sink activity of rice spikelets is predetermined before flowering through carbohydrate supply for developing husks/or pollen grains. Seo and Ota (1983) showed that a limited supply of carbohydrates for spikelets during their development increased the number of malformed spikelets, which became either infertile or half-filled grains. Yamada (1972) and Moriwaki (1999) indicated that excessive plant N and reduced carbohydrate content around meiosis resulted in an abnormal development of pollen grains, which caused spikelet sterilities.
The above review indicates that CGR during the 2-week period has critical effects on final grain yield. Of course, CGR at any stage in rice development influences yield. However, rice grown under intensive nutritional management has a higher LAI, leaf N content, and biomass before the middle of the reproductive period, which often causes growth stagnation in the subsequent stage, presumably because of increased maintenance respiration and mutual shading of leaves. Akita (1989) termed such a phenomenon “over growth”. It is for that reason that CGR during the 2-week period or latter half of reproductive period has critical effects on yield. This implies that maximizing CGR during this period is the important target of rice breeding and managements for maximizing the yield.
Horie et al. (2003a) found that genotypes with higher CGR during the 2-week period had higher radiation use efficiency (RUE) and that higher RUE was associated with higher leaf N concentration, stomatal conductance and hence higher leaf photosynthetic rate under field conditions.
These previous studies suggest that further increase of yield potential necessitates genetic resources for higher leaf photosynthetic rate. The existence of large genotypic difference in stomatal conductance and photosynthetic rate among rice genotypes (Cook and Evans, 1983; Horie et al., 2003a) indicates that there is still a possibility of finding genotypes with higher photosynthetic rate. Further, Uchida et al. (2002) showed a marked transgressive segregation in leaf photosynthetic rate, Rubisco and Rubisco activase contents in progenies of an interspecific hybrid between O. sativa and O. rufipogon. Ku et al. (2000) reported increased leaf photosynthetic rate associated with increased stomatal conductance in transgenic rice in which maize PEPC genes were introduced. By using these genetic resources, photosynthetic rate and yield potential of rice will be increasing at a moderate rate.
Assessment of Crop and Resource Management Technologies
System of rice intensification (SRI)
The system of rice intensification (SRI), originated in Madagascar rice culture and is currently attracting world-wide attentions. As explained by Uphoff and Randriamiharisoa (2002) and Stoop et al. (2002), SRI is a set of principles accompanied by a set of basic management practices:
- Transplanting young seedlings, usually 8 to 12 days old and certainly less than 15 d old and with 2-4 phyllochrons.
- Planting single seedlings rather than several seedlings together in a hill.
- Wide space planting (spacing of 25 × 25 cm or wider).
- Planting quickly and carefully to avoid trauma to seedling roots.
- Daily or intermittent irrigation before panicle initiation (PI) after which a thin layer of water (1-2 cm) is maintained.
- Hand or mechanical weeding, starting 10 days after transplanting and repeated 2-4 times at 10-12 d intervals until the canopy closes.
- Applying nutrients to soil preferably in organic form such as compost or mulch.
- Uphoff and Randriamiharisoa (2002) claimed that double or triple yields over those of conventional rice cultures were attained by SRI in Madagascar. Rafaralahy (2002) reported yields over 15 t ha-1 or even above 20 t ha-1 by SRI in highlands in Madagascar. Criticisms of these reports on SRI were made by Sheehy et al., (2004), Dobermann (2004) and Sinclair and Cassman (2004) for the extraordinary high yields, effectiveness of SRI practices, experimental procedures and publications.
Actually, many of the SRI practice elements have been tried by Japanese farmers including those in yield contest during the 1950s and 1960s. They have also been investigated by rice scientists for their effectiveness over the past 50 years. Since these practice elements cover important areas of rice crop management, we examine their effectiveness in the subsequent section, and here assess briefly SRI as a system.
SRI is a system that require a large amount of labor especially for frequent (3 to 4 times) and intensive weeding. The system of sparse transplanting of young seedlings and unflooding conditions before PI encourages weed damage. In situations where herbicides are not affordable, very labor-intensive weeding is indispensable and, without this, the whole system will be disrupted. In SRI, weeding is practiced by hand and manpower-driven rotary weeder as in Japan, Korea and probably China before the use of herbicides. These weeding methods require hard and patient work. Further, SRI practices require high level of skills in producing a uniform and level puddled field, careful transplanting of small seedlings, avoidance of seedling submergence, replanting gaps, delicate water management to avoid drought damage and weeding without damaging rice plants. Without them, any beneficial effects of SRI are unlikely, and only highly experienced farmers may be able to manage SRI. The intensive labor requirement and high level of skill needed are likely to limit adoption of SRI in Asia. Even in Madagascar, farmers’ adoption of SRI is limited or disadoption is increasing (Moser and Barrett, 2002).
When these requirements for SRI practice are satisfied, however, SRI can be a high-yielding rice system. Many of its components have proven potential to increase rice yield, as described in the subsequent section. In the system of relatively sparse transplanting of one or two seedlings per hill, the application of large amount of compost and intermittent irrigation before PI, rice shows a pattern of relatively slow vegetative growth and progressively more vigorous growth during reproductive development. Such a growth pattern coincide with the hypothesis that maximizing CGR during the 2-week period preceding full heading, under given conditions, is an essential requirement for higher rice yield (Horie, 2001; Horie et al., 2003a).
However, the claim that SRI produced rice yield of 15t ha-1 or more in the highlands of Madagascar (Rafaralahy, 2002) is hard to accept. The Madagascar highlands may have a similar climate to Yunnan where high rice yield is reported (Ying et al., 1998 and also this report) and yield potential there may also be high. However, such high yields seem to exceed the potential yield as simulated by Sheehy et al. (2004) in a model which incorporated a physiologically reasonable value of radiation use efficiency. Clearly more information about experimental procedures, environmental conditions, methods of yield measurements, biomass yield and N uptake is needed, as Sinclair and Cassman (2004) and Dobermann (2004) pointed out.
Also, the claim that SRI could produce double or triple yields over the conventional yields (Uphoff and Randriamiharisoa, 2002) needs careful consideration, because the conventional yields reported were extremely low. They reported that the yields under conventional practice at Morondava, a lowland in Madagascar, were 2.11 and 2.84 t ha-1 for traditional and improved cultivars, respectively, and those at Andriankaja at 1200m in elevation were 2.04 and 3.00 t ha-1, respectively. Since those values are far below than the current world average yield, 5.3 t ha-1 of irrigated rice (Dobermann, 2004), some critical constraints to rice yield were likely to exist in these experimental sites of SRI. The constraints might be either Fe++ toxicity as Dobermann (2004) described, extreme deficiency of nutrients, weed damage or drought. What is necessary would be to identify yield constraints and find ways to minimize effects of constraints one by one. The labor-intensive system of SRI seems not the only way to improve rice yield to the world average.
Elements of SRI practice
As already described, SRI practice elements have been tried and studied for a long period and cover important areas of rice crop management for better yield. We review them here based on previous studies conducted mainly in Japan.
(a) Young seedling transplanting
One of the advantages of transplanting young seedlings lies in higher tolerance to transplanting stresses in younger seedlings than in aged ones. Transplanting causes some stresses to rice, which can be measured as depression in phyllochron development and growth rate after transplanting (Mimoto, 1981; Yamamoto et al., 1995). Under stresses of cool temperatures, high evaporative demand with high temperature, strong wind, high radiation and/or low humidity, transplanted rice loses a portion of its biomass and leaf area, and in the worst case dies, which is called transplanting injury or transplanting shock (Kropff et al., 1995). It was shown that young seedlings recovered from transplanting stress faster than aged seedlings (Sato 1956; Ota 1975; Yamamoto et al., 1995 and 1998), because younger seedlings had higher nitrogen content (Ota, 1975; Yamamoto et al., 1998). At 2-3 phyllochrons, as in SRI, seedlings still have nutrients in the seed endosperm (Fig. 5) and it is known that the endosperm nutrients contribute to faster recovery from transplanting stress (Ota, 1975; Hoshikawa et al., 1995; Yamamoto et al., 1995 and 1998). Yamamoto et al. (1998) reported that, while seedlings with 3.8 phyllochrons transplanted under a mild environment took 5 days to recover in phyllochron development and growth rate, seedlings with 2.2 phyllochrons did so in 2 days, in which phyllochron was counted with incomplete leaf as one.
These earlier studies suggested that young seedlings with less than 4 phyllochrons have higher potential to recover from transplanting stress than those with more than 5 phyllochrons, which are usually used for hand transplanting. Also, quick transplanting from nursery beds to main fields as recommended in SRI may help reduce transplanting injury. Another advantage of early transplanting is because younger seedlings have higher tiller production potential than aged seedlings (Yamamoto et al., 1995 and 1998; Kanda, 1997). Raising rice seedlings in nursery beds for a prolonged period suppresses development of tillers from lower nodes because of early intraspecific competition between plants. Transplanting young seedlings relieves seedlings from this competition. Also, young seedlings are transplanted shallower than aged seedlings and recover more rapidly from transplanting stress as already described. All of these advantages help seedlings develop tillers from lower nodes; Yamamoto et al., (1998) showed that seedlings with 2.0 phyllochrons developed the first tiller from the 2nd node while those with 3.8 phyllochrons did so from the 4th node.
It is well known that rice has the potential to produce one tiller on the nth node when the new leaf on n+3th node emerges (Katayama, 1951). This system is maintained for the relationships between main stem and primary tillers, primary tillers and secondary tillers, and so on. Under this system of synchronous leaf and tiller development in rice, a plant with 13 leaves on the main stem, for instance, has a potential to develop 59 tillers during growth. However, not all of these tillers develop to maturity; some degenerate or get dormant when young and some die later, depending on environmental and nutritional conditions. For a plant with its first tiller bearing on the 2nd node, there is a maximum of 40 tillers, and for a plant with its first tiller on the 4th node, there is a maximum of 18 tillers. Thus, young transplanted seedlings have more potential tillers than older transplanted seedlings.

Fig. 5. Changes with phyllochron in percentage endosperm weight and plant length in rice.
The same relationship also exists between leaf emergence and crown roots emergence: when new leaves emerge on n+3th node on a stem, crown roots on the nth node emerge (Fujii, 1961). Therefore, younger transplanted seedlings also have the potential to develop more roots than older seedlings. Although transplanted younger seedlings have higher potential to develop tillers and roots, the extension of these advantages to higher yield depends on environment and management. The younger the seedling at transplanting, the longer is the time to heading and maturity in the field. In places where growth duration is limited by temperature, as in northern Japan, by availability of water or by the need to plant subsequent crops, young seedlings do not have enough time to express their potential. Young seedlings are also less competitive with weeds and are more susceptible to submergence damage.
(b) Wide space planting with single seedling per hill
The analysis of planting density and yield relationship in rice is an old and new subject of research. Many classical experiments tested planting density from a few hills per m2 to several hundreds hills per m2 (Kanda and Kakizaki, 1956; Yamada et al., 1961; Takeda and Hirota, 1971). Those results generally showed that, with increasing planting density, both biomass and grain yields initially increased and then reached different plateaus . The planting density at which grain yield plateaued was lower than biomass yield. The results of Takeda and Hirota (1971) showed that grain yield was practically unchanged between planting densities from 10 to 100 hills per m2. These results indicate that rice has a wide adaptability to planting density through regulation of panicle numbers, number of spikelets per panicle and grain-filling percentage, depending on environments. Such a response to planting density appears to follow the hypothesis (Kira et al., 1953) that, with increasing planting density, biomass yield reaches a ceiling value that is determined by the supply of resources.
In 1970s in Japan, the rice transplanting system changed from hand transplanting of aged seedlings with phyllochron above 5 to machine transplanting of young seedlings with 3-4 phyllochrons. The high tillering potential of young seedling together with high nitrogen input often caused excessive vegetative growth along with more non-productive tillers, reduced culm diameters of the remaining productive tillers (Kamiji et al., 1993), and lodging increases (Hashikawa, 1996). The new recommendations were to plant seedlings at wide spacing or with fewer plants per hill with reduced basal fertilization(Hashikawa, 1985), and experienced farmers now follow such practices in Japan. Kanda (1997) reported that, transplanting seedlings with 2-3 phyllochrons as in SRI, the optimum planting density to avoid lodging was 13.9 hills m-2, similar to the 16 hills m-2 of SRI practice, but a significantly lower density than the current conventional rate of 20-25 hills m-2 in Japan. These previous studies suggest that, although the effect of planting density on yield is relatively small in rice so long as the density remains within a wide optimal range, sparse transplanting can help reduce non-productive tillers and lodging, especially where young seedlings are transplanted, leading to an efficient and stable rice culture.
The effect of number of plants per hill on yield is similar to the effects of planting density (Yamada et al., 1961). However, San-oh et al. (2004) recently reported a significant effect of single plant per hill on the yield of rice. They compared growth and yield characteristics between 1 plant per hill and 3 plants per hill, at the same number of plants per unit area, both under direct seeding and transplanting conditions. Grain yield at 1 plant per hill exceeded that at 3 plants per hill by 13 % under direct seeding and by 12 % under transplanting. The higher yield at 1 plant per hill was associated with higher CGR after panicle initiation which in turn was associated with higher LAI, larger number of roots per unit area, and higher N and Rubisco contents in the leaf. Interestingly, the plants at 1 plant per hill took up twice as much N than those of 3 plants during the grain-filling period. They ascribed the higher N uptake during grain-filling period to larger number of roots and higher root activity in exudation rate.
Although it is difficult to explain the exact mechanism that produced higher yield at 1 plant per hill, as reported by San-oh et al., (2004), a possible mechanism could be as follows. It is generally accepted that growth of rice roots are supported by substrate supply from lower leaves. Since shading of lower leaves by upper leaves was less severe at 1 plant per hill than that at 3 plants per hill for a prolonged period, more substrate could be supplied to developing roots. Indeed, Tanaka and Arima (1996) reported that, while root weight of densely planted rice reached a maximum before panicle initiation (PI) and then decreased monotonically, that of sparsely planted rice showed two peaks, at PI and at heading. Maintenance of root mass and activity might have contributed more N uptake during the later growth period at 1 plant per hill and contributed for the yield increase, as in the previous report (Okajima, 1966) that N uptake of sparsely planted rice markedly exceeded that of densely planted rice at later growth stages. Anyway, the results of San-oh et al. suggest the necessity of more research on planting density and yield relationships in view of top and root interactions.
(c) Intermittent irrigation and delayed flooding
The concept that rice is not an aquatic plant (Uphoff and Randriamiharisoa, 2002) refects the views of Arashi (1956). Prolonged flooding causes the soil to become increasingly anaerobic with a low redox potential. Depending on soil physico-chemical properties, these conditions cause adverse effects on root development and activity, including reduction in number and diameter of lateral roots (Kawata and Katano, 1977); root respiration (Kido, 1956; Yamada and Ota, 1961) alpha-naphthylamine oxidative power (Hayashi et al., 1960; Yamada and Ota, 1961; Miyasaka, 1975); nutrient uptake (Kido, 1956; Hayashi et al., 1960); root damage and rots due to reduced products of Fe++ and H2S (Baba et al., 1953; Kido, 1956; Ota, 1975; Miyasaka, 1975) in the increasing order of severity. These effects of low redox potential were more pronounced in poorly drained paddy fields rich in clay and organic matters (Kido, 1956; Hayashi et al., 1960; Ota, 1975). The H2S damage also occur in degraded sandy soils along with deficiency of Fe, Mg and Mn (Ota, 1975), which is known as Akiochi (Shioiri, 1943). These adverse effects on roots cause reductions in stomatal conductance (Ishihara et al., 1981), photosynthesis (Koyama et al., 1962), leaf longevity (Iida et al., 1990) and yield (Kido, 1956; Nojima, et al., 1962; Tanaka et al., 1965; Ota, 1975; Iida et al., 1990) to a different degree depending on their severity.
For minimizing those adverse effects of prolonged flooding on rice, and for increasing the yield, various kinds of water management methods have been practiced by farmers and investigated by researchers in Japan for the past 100 years, including midseason drainage, delayed flooding, intermittent irrigation and flush irrigation. Also, the government has promoted installation of drainage infrastructure to paddy field for enhancement of water percolation and drainage. Midseason drainage is a practice to dry fields for a week or so just before panicle initiation. Delayed flooding is to keep the soil moist (about 80% of field capacity) up to panicle initiation and thereafter maintain submerged condition (Arashi, 1956; Tanaka et al., 1965; Heenan and Thompson 1984; Belder et al., 2002), which is essentially the same as the SRI practice. Intermittent irrigation is to repeat 2-4 days moist and 2-4 days submerged condition for part or all of the growth period. The objectives of these water management systems are primarily to supply oxygen to roots and enhance their development and activity, and secondarily to reduce non-productive tillers and suppress stem elongation to avoid lodging when necessary.
These water management systems decrease primary crown root number but increase lateral root number, especially in deeper soil layers (Kawata and Katano, 1977); they decrease percentage of black roots (Iida et al., 1990); increase root respiration rate and alpha-naphthylamine oxidative power (Yamada and Ota, 1961); increase leaf photosynthetic rate (Koyama et al., 1962); and leaf longevity (Iida et al., 1990). However, their effects on yield differ, depending on soil and nutritional conditions. Nojima et al. (1961) and Tanaka et al. (1965) showed that water withdrawal treatments increased yield to about 10% in soils with high N availability but decreased yield more than 10% in soils with low N, because withdrawal of water and irrigation treatments enhanced denitrification and nitrate leaching. Also, Iida et al. (1990) indicated that the positive effect of intermittent irrigation on yield was more pronounced when organic matter was incorporated into the soil.
These previous studies suggest that both intermittent irrigation and delayed flooding reduce the adverse effects of low redox potential on rice roots and allow them to penetrate deeper into the soil, but such effects on rice yield depend on availability of N. In SRI, where a large amount of compost is incorporated, intermittent irrigation before PI may have an effect to promote its decomposition. Application of large amount of compost and intermittent irrigation were also the practices of many of the contest record farmers in Japan during the 1950s and 1960s.
(d) Application of large amount of organic matter
Soil organic matter plays important roles in paddy fields: it provides nutrients source, enhances cation exchange capacity of soil, provides a buffer to drastic changes of root zone environment such as pH, and provides an energy source for soil micro-organisms which also have various activities such as N fixation. Regardless of whether rice is grown by organic farmers or not, input of organic matter in appropriate forms and amounts is essential for sustainable rice farming. This is more so for farmers in many developing countries where chemical fertilizer is too expensive to apply to their fields. Even in high fertilizer input with three or more split N applications, 50% or more of rice uptake-N is shown to be from indigenous source (Japanese Society of Soil Science and Plant Nutrition, 1986). Thus, increasing soil fertility is important for high-yielding rice cultures as mentioned by Cassman et al. (1998). Many of the contest-winning crops that yielded more than 12t ha-1 in 1950s and 1960s in Japan received 10-20t ha-1 or more of compost every year, together with about 100kg N ha-1 as fertilizer (Fuke, 1961). Whether such an amount of composts was necessary is not clear, but at least it gave no negative effect.
Aerobic rice
Aerobic rice is defined as high-yielding rice grown in nonpuddled and nonflooded aerobic soil (Bouman, 2001). It is usually grown under supplementary irrigation and with fertilizer inputs (Wang et al., 2002). A similar system was studied and practiced in Japan during the 1950s and 1960s (Umino et al., 1959; Hasegawa et al., 1960) where lowland cultivars were grown instead of upland cultivars. Much attention has been paid to aerobic rice in China since mid-1980s where water conservation is becoming an important issue.
Specific cultivars for aerobic rice cultures are being bred in China (Wang et al., 2002; Yang et al., 2002). Wang et al. (2002) showed that yields of aerobic rice cultivars grown in aerobic soil nearly doubled those of traditional upland cultivars, but were 20-30% lower than those of lowland cultivars grown under flooded conditions. They further showed that the water use of aerobic rice culture was about 60% less than that of lowland rice. Similar results were also reported by Hasegawa et al. (1960) and Yang et al. (2002). To further save water in aerobic rice, attempts were made to cover the soil surface with plastic film, a practice called ground cover rice production system (GCRPS; Lin et al., 2002).
As those results in China and Japan showed aerobic rice provides significant water saving compared with lowland rice. Productivity of traditional upland rice will largely be improved by replacing it by aerobic rice with irrigation. However, as already noted for water management of SRI, drying paddy soil increases denitrification and nitrate leaching. Indeed, a drastic reduction in plant uptake of fertilized N was reported in aerobic rice (Lin et al., 2002). Thus, aerobic rice is a system that can save water at the expense of N. Adequate N management including organic matter applications or use of controlled release fertilizer is obviously necessary for sustainable and increased aerobic rice production.
Improved nitrogen management
As shown in Fig. 3, the rate of N fertilization in Shiga prefecture began to decrease in 1980s and it now averages around 70 kg ha-1 as compared with over 100 kg ha-1 in late 1970s. Nevertheless, the average grain yield was maintained or it rather increased moderately in the same period and thus improvement of agronomic efficiency of fertilized N (AE) (Novoa and Loomis, 1981) is evident in this region. The R-DYA (i.e., the average yielding ability of cultivars) also decreased gradually during this period (Fig. 3). Therefore, the increase of AE should be attributed to improved crop management.

Fig. 6. Typical values of recovery efficiency and agronomic efficiency (d(grain yield)/(N fertlized)) of various N fertilization methods for irrigated rice in Japan and Philippines. Single/split appl.: single dose application at transplanting or split application at more than three times of urea or ammonium sulfate2-6). Local. appl.: Urea tablet deep placed at 14d after transplanting 1) or band placement of urea at transplanting 3). SPAD based split: urea top-dressed whenever the chlorophyll meter value of the topmost fully expanded leaf fell below 355). CRU: controlled release urea (polyolefin-coated urea) incorporated in the soil at transplanting3, 7-9). CRU local. 1: CRU locally placed at transplanting or direct-seeding10, 11). CRU local. 2: CRU applied to the nursery bed and applied to the paddy soil being attached to the seedling at transplanting10, 12,13).
Recovery efficiencies are based only on the 15N method. The agronomic efficiency with * is the estimated value from recovery efficiency, multiplying it to an assumed value of physiological efficiency (d(grain yield)/d(N uptake)); 55 kg kg-1.
Reproduced from 1)Balasubramanian et al., 1999; 2)Okamoto et al., 1986; 3)Shibahara et al., 1992; 4)Peng et al., 1996a; 5)Peng et al., 1996b; 6)Shiraiwa et al., 1999; 7)Ueno et al., 1991; 8)Sato et al. 1993; 9)Sato et al., 1997; 10)Kaneta et al., 1994; 11)Ando et al., 2000; 12)Kaneta and Tsuchiya, 1997a; 13)Kaneta and Tsuchiya, 1997b.
Technological development of N application is illustrated in Fig. 6. The recovery efficiency (RE) is below 30% when ammonium sulfate or urea is applied at transplanting. Numerous studies have shown that the RE of N can be improved by split application and it is most efficient when N-dose distribution is designed to match N demand by the growing crop, typically monitored with leaf color charts (Matsuzaki et al., 1980) or the chlorophyll meter (SPAD) (Peng et al. 1996). Reflecting such a concept of N application, the extension authority in Japan changed the recommendation of N fertilization in early 1980s from 70% basal and early dressings to one with decreased early dressings (30%) and increased later dressings (70%) and a reduced total amount. The new recommendation has been widely accepted among farmers (Shibahara, 1992). Localized application of N at transplanting, which has been mechanized in Japan with a band fertilizer-locator attached to the planting machine, performed higher RE than 40% (Shibahara et al., 1992). This local placement method, established in mid 1980s, is presently adopted by around 40% of rice farmers in Shiga. Further enhancement of RE was achieved by slow release fertilizers, especially by olefin-resin coated urea, generally called the controlled release urea (CRU). The combination of CRU and localized application techniques achieves RE as high as 80% (Kaneta et al.; 1994, Ando et al., 2000; Kaneta and Tsuchiya, 1997a, b). This kind of slow-release fertilizer also has been adoped in Shiga on 20% of farms. Although the costs for CRU and mechanized local placement are high compared with conventional techniques, it has been offset in Japan by the reduced labor cost. The AE of fertilization methods has been enhanced with change in RE as shown in Fig. 6. Thus, new N application methods have certainly contributed to yield maintenance/increase with decreasing amount of N application in Shiga.
In addition, N release of CRU is highly predictable in relation to soil temperature and this may play a role in higher-yielding production. It is because the N release pattern of CRU matches the changing crop demand for N and, at least in the temperate region, the highest temperature season coincides with the important growth stage for yield formation, especially the late reproductive period (Horie, 2001).
Production Technologies toward Increased Yield in Irrigated Rice
The above review showed that yield of the best Chinese hybrid, exceeded yields of the best inbred cultivars by about 10%. This and the slackened yield increase of inbred cultivars indicated that increase in rice yield potential has been moderate in these decades. Although attempts are being made to introduce C4 photosynthesis genes into rice (Ku et al., 2000), it is unclear when and to what extent such a transgenic rice will contribute for the increase in rice yield potential. Under such a situation, the increase in rice yield potential will continue to be moderate, and it alone may not enough to fulfill the increasing rice demands in the coming decades.
It was shown that the contest record yields in Japan in 1950s and 1960s nearly tripled the average yield at that time and doubled the current average yield. This implies that the current crop and resource management technology is exploiting only a part of large yield potential of rice. Although application of system of rice intensification (SRI; Uphoff and Randriamiharisoa, 2002) to Asia in its form is difficult for its high labor requirement, its practice elements reminded us important points toward crop and resource management for increased yield in rice.
The practice of transplanting one or two seedlings per hill has a potential to increase rice yield, through reducing transplanting stress or injury and increasing tiller and root number on lower nodes. This potential can most fully be realized by direct seeding rather than transplanting. It is true that direct-seeding system still have problems in seedling establishment, weed control and lodging, but, when those problems are overcome, direct-seeded rice shows higher yield than transplanted rice (Yoshinaga et al., 2002; Tabbal et al., 2002; San-oh et al., 2004). New technologies have been developed to overcome these problems of direct-seeding rice system: coating pre-germinated seeds with calcium peroxide to facilitate seedling establishment in anaerobic conditions (Ota and Nakayama, 1970); breeding more lodging-tolerant genotypes; effective methods of herbicide use and mechanization of direct-seeding on wet or submerged soils (Yoshinaga et al., 2002). With further improvement of those technologies, direct-seeding can be a high-yielding rice production system. However, it should be kept in mind that transplanted rice has the advantages that it is highly competitive with weeds, gives higher yields where growth duration is limited due to cool climate or availability of water; and enables multiple cropping of 2 or 3 crops per year. In places where these limitations do not apply, direct seeding system can be a labor-efficient and high-yielding rice production system.
The combined practice of organic matter incorporation and intermittent irrigation also has a potential to increase rice yield, when it is combined with effective use of fertilizers. This, and deep plowing and installation of drainage facility are considered the major practices that enabled extremely high rice yield of the contest farmers in 1950s and 1960s. In such a system, rice develops a deeper root system and maintains its activity to later growth stages as discussed already, which might facilitate uptake of slowly mineralized N at later stages. There is also suggestion that high-yielded rice had deeper root system (Kawata et al., 1978; Morita et al., 1988), but the exact mechanism for this is the subject of further studies.
Improved N management has not only contributed to increased rice yield also to more efficient use of fertilizers. Controlled release fertilizer is increasing yield with reduced N application, through its ability to supply N that closely matches rice demand. Currently, this fertilizer is too costly in most rice producing countries. However, there are many N management options that enable similar outcomes as controlled release fertilizers. Among all, N management based on crop diagnosis as indicated by Peng et al., (1996) may be most appropriate. Instead of a costly chlorophyll meter, leaf color charts (Matsuzaki et al., 1980) are available with modest cost. Diagnosis with use of this chart and careful field observation of rice growth and development will enable rice yield increase through efficient use of fertilizer.
As described above, there are many opportunities to increase rice yield by improved crop and resource managements. It is important to integrate these technological elements into a system that is suited for and adaptable to regional environments. This, together with moderate increases in yield potential of genotypes will enable the resumption of rice yield increases to a comparable level to that of the 1980s.
References
Akita, S. 1989. Improving yield potential in tropical rice. In: Progress in Irrigated Rice Research. IRRI, Los Baños, Philippines. 41-73.
Ando, H., Kakuda, K., Nakamura, M. and Yokoto, K. 2000. Yield of no-tillage direct-seeded lowland rice as influenced by different sources and application methods of fertilizer nitrogen. Soil Sci. Plant Nutr. 46: 105-115.
Arashi, K. 1956. Chapter 74. Irrigation and drainage. In Y. Togari and T. Matsuo eds., Rice Culture 3., Asakura Shoten, Tokyo. 191-204 (in Japanese).
Baba, I, Takahashi, Y. and Iwata, I. 1953. Studies on the nutrition of the rice plant with reference to helminthosporium leaf spot (preliminary report), V. The plant growth and susceptibility to the disease as affected by the addition of iron to culture solution. Proc. Crop Sci. Soc. Jpn. 22: 41-42 (in Japanese with English summary).
Balasubramanian, V., Morales, A.C., Cruz, R.T. and Abdulrachman, S. 1999. On-farm adaptation of knowledge-intensive nitrogen management technologies for rice systems. Nutr. Cycl. Agroecosys. 53: 59-69.
Belder, P., Bouman, B.A.M., Spiertz, J.H.J., Lu, G. and Quilang, E.J.P. 2002. Water use of alternately submerged and nonsubmerged irrigated lowland rice. In B.A.M. Bouman et al. eds., Water-Wise Rice Production. IRRI, Los Baños, Philippines. 51-61.
Bouman, B.A.M. 2001. Water-efficient management strategies in rice production. Int. Rice. Res. Notes 16: 17-22.
Cassman, K.G., Peng, S., Olk, D.C., Ladha, J.K., Reichardt, W., Dobermann, A. and Singh, U. 1998. Opportunities for increased nitrogen-use efficiency from improved resource management in irrigated rice systems. Field Crops Research 56: 7-39.
Cock, J.H. and Yoshida, S. 1972. Accumulation of 14C labelled carbohydrate before flowering and the subsequent redistribution in the rice plant. Proc. Crop Sci. Soc. Japan 41: 226-234.
Cook, M.G. and Evans, L.T. 1983. Some physiological aspects of the domestication and improvement of rice (Oryza spp.). Field Crops Res. 6: 219-238.
Dobermann, A. 2004. A critical assessment of the system of rice intensification (SRI). Agricultural Systems 79: 261-281.
Feught, C. and Hofner, W. 1985. Influence of growth regulators and nitrogen on the number of endosperm cells in wheat grains from different positions on the ear. J. Plant Physiol. 117: 469-478.
Feyerherm, A.M., Paulsen, G.M. and Sebaugh, J.L. 1984. Contribution of genetic improvement to recent wheat yield increases in the USA. Agron. J. 76: 985-990.
Fujii, Y. 1961. Studies on the regularity of root growth in rice and wheat plants. Agric. Bull. Saga Univ. 12: 1-117 (in Japanese with English summary).
Fukai, S. and Cooper, M. 1995. Development of drought-resistant cultivars using physio-morphological traits in rice. Field Crops Res. 40: 67-86.
Fuke, Y. 1961. Analysis of technological effects on rice contest yields in Japan -Hokuriku district-. Tokyo Agric. Univ., Tokyo. 1-74 (in Japanese).
Hasegawa, H., Takemura, G. and Nakayama, K. 1960. Paddy rice growing in upland field - The adaptability to row spacing and sowing date. Proc. Crop Sci. Soc. Japan 29: 19-22 (in Japanese with English summary).
Hasegawa, T. and Horie, T. 1995. Evaluation of relative contributions of cultivar and crop management to regional yield increase of rice. -For Kinki district from 1960 to 1980-. Agriculture and Horticulture. 70: 233-238 (in Japanese).
Hasegawa, T., Horie, T. and Yandell, B.S. 1991. Improvement of yielding ability in Japonica rice cultivars and its impact on regional yield increase in Kinki district, Japan. Agricultural Systems 35: 173-187.
Hasegawa, T., Koroda, Y., Seligman, N.G. and Horie, T. 1994. Response of spikelet number to plant nitrogen concentration and dry weight in paddy rice. Agron. J. 86: 673-676.
Hashikawa, U. 1985. Principal Techniques for Rice Culture. Nobunkyo, Tokyo. 1-238 (in Japanese)..
Hashikawa, U. 1996. Low input rice culture. Fumin-Kyokai, Osaka. 1-366. (in Japanese).
Hayashi, M. Hashizume, A., Shinohara, S. and Igarashi, G. 1960. Studies on the influence of water-percolation in rice plants during ripening period in ill-drained paddy field. Proc. Crop Sci. Soc. Jpn. 29: 43-46 (in Japanese with English summary).
Hayashi, M. 1961. The Early Planting Rice Culture. Yokendo Ltd., Tokyo. 1-306. (in Japanese).
Heenan, D.P. and Thompson, J.A. 1984. Growth, grain yield and water use of rice grown under restricted water supply in New South Wales. Aust. J. Exp. Agric. 24: 104-109.
Horie, T. 2001. Increasing yield potential in irrigated rice: breaking the barrier. In S. Peng and B.Hardy eds. Rice Research for Food Security and Poverty Alleviation. Proc. Int. Rice Res. Conf., 31 Mar. - 3 Apr. 2000, Los Baños, Philippines, IRRI. 3-25.
Horie, T., Ohnishi, M., Angus, J.F., Lewin, L.G., Tsukaguchi, T. and Matano, T. 1997. Physiological characteristics of high-yielding rice inferred from cross-location experiments. Field Crops Res. 52: 55-67.
Horie, T., Lubis, I., Takai, T., Ohsumi, A., Kuwasaki, K., Katsura, K. and Nii, A. 2003a. Physiological traits associated with high yield potential in rice. In T.W. Mew et al. eds. Rice Science: Innovations and Impact for Livelihood, IRRI, Los Baños, Philippines. 117-145.
Horie, T., Yoshida, H., Shiraiwa, T., Nakagawa, H., Kuroda, E., Sasaki, T., Hagiwara, M., Kobata, T., Ohnishi, M. and Kobayashi, K. 2003b. Analysis of genotype by environment interaction in yield formation processes of rice grown under a wide environmental range in Asia. 1. Asian Rice Network (ARICENET) research and preliminary results. Jpn. J. Crop Sci. 72 (Extra issue 2): 88-89 (in Japanese).
Hoshikawa, K. 1967. Studies on the development of endosterm in rice 3. Observation on the cell division. Proc. Crop Sci. Soc. Japan. 36: 210-215.
Hoshikawa, K., Sasaki, R. and Hasebe, K. 1995. Development and rooting capacity of rice nurseling seedlings grown under different raising conditions. Jpn. J. Crop Sci. 64: 328-332.
Hossain M. 1999. Longterm perspective of the demand-supply balances for rice in Asia: Implications for technological challenge. In T. Horie et al. eds. Proc. Int. Symp. "World Food Security", Crop Sci. Soc. Jpn., Tokyo. 31-39.
Iida, S., Shinmura, Y., Uemori, A. and Kuzuna, K. 1990. Influence of irrigation management at the middle growing stage on rice plant growth and yield. Jpn. J. Crop Sci. 59: 413-418 (in Japanese with English abstract).
IRRI 1997. Rice Almanac. International Rice Research Institute, Los Baños, Philippines. 1-181.
Ishihara, K., Hirasawa, T., Iida, O. and Kumura, M. 1981. Diurnal course of transpiration rate, stomatal conductance, xylem water potential and leaf water potential in the rice plants under the different growth conditions. Jpn. J. Crop Sci. 50: 25-37 (in Japanese with English abstract).
Japanese Society of Soil Science and Plant Nutrition 1986. Nitrogen Mineralization in Paddy Soil and Fertilization. Hakuyusha, Tokyo. 1-181 (in Japanese).
Kamiji, Y., Hayashi, S. and Horie, T. 1993. Influences of nitrogen nutrient and solar radiation in the canopy on length of lower internodes of rice. Jpn. J. Crop Sci. 62: 164-171 (in Japanese with English summary).
Kanda, Y. 1997. Rice culture with sparse planting of nurseling seedling. In Nogyo Gijutsu Taikei, Field Crop 2, Suppl. Issue 19, Nobunkyo, Tokyo. 522.2-522.9 (in Japanese).
Kanda, K. and Kakizaki, Y. 1956. Population ecological studies on planting density in rice. Proc. Crop Sci. Soc. Japan 25: 59 (in Japanese).
Kaneta, Y. and Tsutiya, K. 1997a. Relationship between variation of weather and recovery rate of rice plants in non-tillage culture by single application of fertilizer in nursery boxes. Jpn. J. Soil Sci. Plant Nutr. 68: 112-115.
Kaneta, Y. and Tsutiya, K. 1997b. Effects of single application of fertilizer in nursery boxes on growth and nitrogen uptake of rice in wet paddy fields with rice straw plowed. Jpn. J. Soil Sci. Plant Nutr. 68: 185-188.
Kaneta, Y., Awasaki, H. and Murai, Y. 1994. The non-tillage rice culture by single application of fertilizer in a nursery box with controlled-release fertilizer. Jpn. J. Soil Sci. Plant Nutr. 65: 385-391.
Katayama, T. 1951. A Study on Tillering Systems in Rice and Wheat. Yokendo Ltd., Tokyo. (in Japanese).
Kawata, S. 1976. Japanese Crop Cultivation Technologies. Yokendo Ltd., Tokyo. 1-428. (in Japanese).
Kawata, S. and Katano, M. 1977. Effect of water management of paddy fields on the direction of crown root growth and the lateral root formation of rice plants. Jpn. J. Crop Sci. 46: 543-557 (in Japanese with English summary).
Kawata, S., Soejima, M. and Yamazaki, K. 1978. The superficial root formation and yield of hulled rice. Jpn. J. Crop Sci. 47: 617-628 (in Japanese with English summary).
Khush, G.S. and Peng, S. 1996. Breaking the yield frontier of rice. In M.P. Reynolds et al. eds., Increasing Yield Potential in Wheat: breaking the barriers. Proc. of a workshop held 26-28 March, 1996 in Cindad Obregon, Sonora, International Maize and Wheat Improvement Center, Mexico.
Kido, M. 1956. Ill-drained and well drained paddies. In Y. Togari and T. Matsuo eds., Rice Culture 3., Asakura Shoten, Tokyo. 191-204 (in Japanese).
Kira, T., Ogawa, H. and Sakazaki, N. 1953. Intraspecific competition among higher plants I. Competition-yield-density interrelationship in regularly dispersed population. J. Inst. Poly. Osaka City Univ. D4: 1-16.
Koyama, T., Miyasaka, A. and Eguchi, K. 1962. Studies on water management in the ill-drained paddy field. VII. The effect of the surface drainage on the photosynthetic activity in the rice plant. Proc. Crop Sci. Soc. Jpn. 30: 143-145 (in Japanese with English summary).
Kropff, M.J. Mathews, R.B., van Laar, H.H. and ten Berg, H.F.M. 1995. The rice model Oryza 1 and its testing. In Mathews et al. eds. Modeling the impact of climate change on rice production in Asia. IRRI, Los Baños, Philippines. 27-50.
Ku, M.S.B., Cho, D., Ranade, U., Shu, T.P., Li, X., Jiao, D-M, Ehleringer, J. and Miyao, M. 2000. Photosynthetic performance of transgenic rice plants over expressing maize C4 photosynthtic enzymes. In Sheehy et al. eds. Redesigning Rice Photosynthesis to Increase Yield. IRRI, Los Baños, Philippines. 193-204.
Lin, S., Dittert, K., Tao, H., Kreye, C., Xu, T., Shen, Q., Fan, X. and Sattelmacher, B. 2002. The ground-cover rice production system (GCRPS): a successful new approach to save water and increase nitrogen fertilizer efficiency? In B.A.M. Bouman et al. eds., Water-Wise Rice Production. IRRI, Los-Baños, Philippines. 187-196.
Matsushima, M. 1957. Analysis of developmental factors determining yield and yield prediction in lowland rice. Bull. Natl. Inst. Agr. Sci. Ser. A 5: 1-271 (in Japanese with English summary).
Matsuzaki, A., Kariya, K., Machida, H. and Tsunoda, K. 1980. Studies on the growth control and the nutritional diagnosis in rice plant. 1. The nutritional diagnosis based on the leaf color and the estimation of the number of spikelets per unit area. Jpn. J. Crop Sci. 49: 439-444 (in Japanese with English summary).
Mimoto, H. 1981. Studies on rooting of rice 5. Problems associated in comparing rooting ability among seedlings with different age. Jpn. J. Crop Sci. 50 (Extra issue 2): 1-2 (in Japanese).
Miyasaka, A. 1975. Crop Management and Production Ecology. In Nogyo Gijutsu Taikei, Field Crop 1, Nobunkyo, Tokyo. 277-291 (in Japanese).
Morita, S., Suga, T. and Yamazaki, K. 1988. Relationship between root length density and yield in rice plants. Jpn. J. Crop Sci. 57: 438-443 (in Japanese with English abstract).
Moriwaki, T. 1999. On the ideal culm elongation in rice plants (3). Nogyogijutsu 54: 133-138 (in Japanese).
Moser, C.M. and Barrett, C.B. 2002. The system of rice intensification in practice: explaining low farmer adoption and high disadoption. In B.A.M. Bouman et al. eds., Water-Wise Rice Production. IRRI, Los Baños, Philippines. 103-118.
Nakamura, T., Matsunaga, D. and Hoshikawa, K. 1992. Relationship between initial growth of spikelet, and fertility and number of endosperm cells in rice. Jpn. J. Crop Sci. 61 (Extra issue 2): 129-130 (in Japanese).
Nojima, K., Tanaka, I. and Uemura, Y. 1961. Influence of drainage on the growth of rice plant in rice field. I) Effect of drainage of surface water on the growth, yield and nutrient uptake of rice plant. Proc. Crop Sci. Soc. Jpn. 30: 321-324 (in Japanese with English summary).
Nojima, K., Tanaka, I. and Uemura, Y. 1962. In Nogyo Gijutsu Taikei, Field Crop 1, Nobunkyo, Tokyo. 259-275 (in Japanese).
Novoa R. and Loomis R. S. 1981. Nitrogen and plant production. Plant and Soil 58: 177-204.
Okajima, H. 1966. Rice development and environment. In Agr. Res. Inst. Tohoku Univ. ed. Theory and direction of development of rice production technology. Tohoku University, Sendai, Japan. 131-148 (in Japanese).
Okamoto, M., Ohashi, K., Hasegawa, K. and Nishikawa, Y. 1986. Effect of fertilizer application methods for machine-transplanted rice on dry matter production and balance of nitrogen. 1. On the growing process and the nitrogen uptake. Bull. Shiga Pref. Agric. Exp. Stn. 27: 1-16 (in Japanese).
Ota, Y. 1975. Site condition and rice crop ecology. In Nogyo Gijutsu Taikei, Field Crop 1, Nobunkyo, Tokyo. 259-275 (in Japanese).
Ota, Y. and Nakayama, M. 1970. Effect of seed coating with calcium peroxide on germination under submerged condition on rice plant. Proc. Crop Sci. Soc. Jpn. 39: 535-536 (in Japanese with English summary).
Pantuwan, G.., Fukai, S., Cooper, M., Rajatasereekul, S. and O'toole, J.C. 2002. Yield response of rice (Oryza sativa L.) genotypes to drought under rainfed lowlands - Part 2. Selection of drought resistant genotypes. Field Crops Res. 73: 169-180.
Peng, S., Khush, G.S. and Cassman, K.G. 1994. Evolution of the new plant ideotype for increased yield potential. In Cassman, K.G. ed., Breaking the Yield Barrier. Proc. of a workshop on Rice Yield Potential in Favorable Environments. IRRI, Los Baños, Philippines. 5-20.
Peng. S., Garcia, F.V., Gines, H.C., Laza, R.C., Samson, M.I., Sanico, A.L., Visperas, R.M., and Cassman, K.G. 1996a. Nitrogen use efficiency of irrigated tropical rice established by broadcast wet-seeding and transplanting. Fert. Res. 45: 123-134.
Peng, S., Garcia, F.V., Laza, R.C., Sanico, A.L., Visperas, R.M. and Cassman, K.G. 1996b. Increased N-use efficiency using a chlorophyll meter on high-yielding irrigated rice. Field Crops Res. 47: 243-252.
Peng, S., Cassman, K.G., Virmani, S.S., Sheehy, J. and Khush, G.S. 1999. Yield potential trends of tropical rice since the release of IR8 and the challenge of increasing rice yield potential. Crop Sci. 39: 1552-1559.
Radley, M. 1978. Factors affecting grain enlargement in wheat. J. Exp. Bot. 29: 919-934.
Rafaralahy, S. 2002. An NGO perspective on SRI and its origins in Madagascar. In N. Uphoff et al. eds., Assessment of the System for Rice Intensification (SRI). Proc. Int. Conf. Sanya, China, April 1-4, 2002. Cornell Int. Inst. Food Agric. and Develop., Ithaca, NY. 17-22.
San-oh, Y., Mano, Y., Ookawa, T. and Hirasawa, T. 2004. Comparison of dry matter production and associated characteristics between direct-sown and transplanted rice plants in a submerged paddy field and relationships to planting pattern. Field Crops Res. 87: 43-58.
Sato, K. 1956. Root development. In Y. Togari and T. Matsuo eds., Rice Culture 2, Asakura Shoten, Tokyo. 15-27 (in Japanese).
Sato, T., Shibuya, K., Saigusa, M. and Abe, T. 1993. Single basal application of total nitrogen fertilizer with controlled-release coated urea on no-tilled rice culture. Jpn. J. Crop Sci. 62: 408-413 (in Japanese with English abstract).
Sato, T., Shibuya, K. and Saigusa, M. 1997. A single basal application of controlled-release coated urea to rice cultures using pot seedlings in a cool region. Jpn. J. Crop Sci. 66: 11-16 (in Japanese with English abstract).
Seo, S.W. and Ota, Y. 1983. Role of the hull in the ripening of rice plant. Bull. Natl. Inst. Agr. Sci. Ser. D 35: 33-102.
Sheehy, J.E., Peng, S., Dobermann, A., Mitchell, P.L., Ferrer, A., Yang, J., Zou, Y., Zhong, X. and Huang, J. 2004. Fantastic yields in the systems of rice intensification: fact or fallacy? Field Crops Res. 88: 1-8.
Shi, Q., Zeng, X., Li, M., Tan, X. and Xu, F. 2002. Effect of different water management practice on rice growth. In B.A.M. Bouman et al. eds., Water-Wise Rice Production. IRRI, Los Baños, Philippines. 3-13.
Shibahara, F., Tsuji, T. and Nishimura, M. 1992. Improvement on nitrogen application efficiency to paddy rice and reduction in the outflow of fertilizer elements by using the coated urea fertilizer. Bull. Shiga Pref. Agric. Exp. Stn. 33: 17-29 (in Japanese with English summary).
Shioiri, M. 1943. Chemistry of Paddy Soil. Extension Report for Improvement of Fertilization 5 (In Japanese).
Shiraiwa, T., Hashikawa, U. and Liu, Y. 1999. Productivity of rice cultivars in reference to nitrogen utilization and its efficiency. -N use for yield formation in high-yielding F1 hybrid rices-. Proc. Int. Symp. "World Food Security", Crop Sci. Soc. Jpn. Tokyo. 294-295.
Shoji, S. and Kanno H. 1994. Use of polyolefin-coated fertilizers for increasing fertilizer efficiency and reducing nitrate leaching and nitrous oxide emissions. Fert. Res. 39: 147-152.
Sinclair, T.R. and Cassman, K.G. 2004. Agronomic UFOs. Field Crops Res. 88: 9-10.
Singh, B.K. and Jenner, C.F. 1984. Factors controlling endosperm cell number and grain dry weight in wheat: Effect of shading on intact plants and of variation in nutritional supply to detached cultured ears. Aust. J. Plant Physiol. 11: 151-163.
Stoop, W.A., Uphoff, N. and Kassam, A. 2002. A review of agricultural research raised by the system of rice intensification (SRI) from Madagascar: opportunities for improving farming systems for resource-poor farmers. Agricultural Systems 71: 249-274.
Sumi, A., Hakoyama, S., Weng, J.H., Agata, W. and Takeda, T. 1996. Analysis of plant characteristics determining ear weight increase during the ripening period in rice (Oryza sativa L.) II. The role of the reserved carbohydrate at heading stage upon the receptive efficiency of assimilation products in spikelets. Jpn. J. Crop Sci. 65: 214-221 (in Japanese with English abstract).
Tabbal, D.F., Bouman, B.A.M., Bhuiyan, S.I., Sibayan, E.B. and Sattar, M.A. 2002. On farm strategy for reducing water input in irrigated rice; Case studies in the Philippines. Agric. Water Manag. 56: 93-112.
Takeda, T. and Hirota, O. 1971. Relationship between spacing and grain yield of rice plant. Proc. Crop Sci. Soc. Japan 40: 381-385.
Takeda, T., Weng, J.H. and Agata, W. 1980. The role of reserved carbohydrate at heading in ear weight increase during the grain-filling of rice. Jpn. J. Crop Sci. 49 (Extra issue 2): 75-76 (in Japanese).
Tanaka, I., Nojima, K. and Uemura, Y. 1965. Influence of drainage on the growth of rice plant in rice field. 3. The influence of irrigation management on the nutrient uptake, nitrogen content in soil. Proc. Crop Sci. Soc. Jpn. 33: 335-343 (in Japanese with English summary).
Tanaka, N. and Arima, S. 1996. Effects of planting density on growth and distribution of roots in rice plants (Oryza sativa L.). Jpn. J. Crop Sci. 65: 71-76 (in Japanese with English abstract).
Tsukaguchi, T., Horie, T. and Ohnishi, M. 1996. Filling percentage of rice spikelets as affected by availability of non-structural carbohydrates at the initial phase of grain filling. Jpn. J. Crop Sci. 63: 445-452 (in Japanese with English summary).
Uchida, N., Matsumoto, C. and Ishii, T. 2002. Genes from wild rice enhance rice photosynthesis. Jpn. J. Crop Sci. 71 (Extra issue 1): 294-295 (in Japanese).
Ueno, M., Kumagai, K., Togasi, M. and Tanaka, N. 1991. Basal application technique of whole nitrogen using slow-release coated fertilizer based on forecasting of soil nitrogen mineralization amount. Jpn. J. Soil Sci. Plant Nutr. 62: 647-653.
Umino, S., Iizuka, S., Nonaka, F. and Yuyama, H. 1959. Studies on methods of lowland rice culture on upland fields under irrigation. I. Applicability of lowland rice variety to the cultivation on upland under irrigation. II. Differences in the growth and physiological characteristics between lowland rice and upland rice cultivated on upland field under irrigation. Proc. Crop Sci. Soc. Japan 27: 349-353 (in Japanese with English summary).
Uphoff, N. and Randriamiharisoa 2002. Reducing water use in irrigated rice production with the Madagascar System of Rice Intensification (SRI). In B.A.M. Bouman et al. eds., Water-Wise Rice Production. IRRI, Los Baños, Philippines. 71-87.
Virmani, S.S. 2001. Opportunities and challenges of developing and using hybrid rice technology in the tropics. In S. Peng and B. Hardy eds. Rice Research for Food Security and Poverty Alleviation. IRRI, Los Baños, Philippines. 151-166.
Wada, G. 1969. The effect of nitrogenous nutrition on the yield-determining process of rice plant. Bull. Natl. Inst. Agric. Sci. Ser. A 16: 27-167 (in Japanese with English summary).
Wang, H. Bouman, B.A.M., Zhao, D., Wang, C. and Moya, P.F. 2002. Aerobic rice in northern China: Opportunities and challenges. In B.A.M. Bouman et al. eds., Water-Wise Rice Production. IRRI, Los Baños, Phillipines. 143-154.
Yamada, N. 1972. Studies on the developmental physiology in rice pollen I. The metabolic patterns connected with the structural changes in developing pollen. Proc. Crop Sci. Soc. Japan 41: 320-334.
Yamada, N. and Ota, Y. 1961. Effect of water percolation on physiological activity of rice root. Proc. Crop Sci. Soc. Jpn. 29: 404-408 (in Japanese with English summary).
Yamada, N., Ota, Y. and Nakamura, H. 1961. Ecological effects of planting density on growth of rice plant. Proc. Crop Sci. Soc. Japan 29: 329-333.
Yamamoto, Y., Ikejiri, A. and Nitta, Y. 1995. Characteristics of rooting and leaf emergence rate, early growth and heading date of rice seedlings with different plant age in leaf number. Jpn. J. Crop Sci. 64: 556-564 (in Japanese with English abstract).
Yamamoto, Y., Ikejiri, A. and Nitta, Y. 1998. Characteristics of taking root of rice nurseling seedling in relation to the changes of some inoganic and organic constituents after transplanting. Jpn. J. Crop Sci. 67: 20-25 (in Japanese with English abstract).
Yang, X., Wang, H. Wang, Z., Zhao, J., Chen, B. and Bouman, B.A.M. 2002. Yield of aerobic rice (HanDao) under different water regimes in North China. In B.A.M. Bouman et al. eds. Water-Wise Rice Production. IRRI, Los Baños, Philippines. 155-164.
Ying, J.Y., Peng, S., He, Q., Yang, H., Yang, C., Visperas, R.M. and Cassman, K.G. 1998. Comparison of high-yield rice in tropical and subtropical environments I. Determinants of grain and dry matter yields. Field Crops Res. 57: 71-84.
Yoshinaga, S., Wakimoto, K., Tasaka, K., Matsushima, K., Togashi, T. and Shimotsubo, K. 2001. Growth characteristics of submerged hill-seeded rice (Oryza sativa L.) in warmer regions of Japan. -Differences of growth as compared with broadcast-seeded rice-. Jpn. J. Crop Sci. 70: 541-547 (in Japanese with English abstract).
Yuan, L. 1994. Increasing yield potential in rice by exploitation of heterosis. In S.S. Virmani ed., Hybrid Rice Technology: New developments and futre prospects. International Rice Research Institute, Los Baños, Philippines. 1-6.
Yuan, L. 2001. Breeding of super hybrid rice. In S. Peng and B. Hardy eds., Rice Research for Food Security and Poverty Alleviation. Proc. Int. Rice Res. Conf., 31 March - 3 April 2000, Los Baños, Philippines. 143-149.
Zou, J., Lu, C., Wang, C., Zheng, K., Zong, S., Zhao, L. and Sun, Y. 2003. Breeding of two-line hybrid rice variety ‘Liangyoupeijiu’ and preliminary studies on its cultivation characters. Scientia Agricultura

Search Site
- Crop Science
- Help/Feedback
- Contact us
- Sitemap