



Linking physiological and genetic analyses of the control of leaf growth under fluctuating environmental conditions
INRA - ENSAM. Laboratoire d'Ecophysiologie des Plantes sous Stress Environnementaux. Montpellier, France. tardieu@ensam.inra.fr . http://www.montpellier.inra.fr/lepse
The respective importance of the mechanisms which drive plant responses to environmental conditions are a function of climatic scenarios, with high year-to-year and site-to-site variabilities. Modelling is therefore an important tool for designing 'stress tolerant' plants. After reviewing mechanisms involved in the response of leaf growth to water deficit, we conclude that gene regulatory networks are not feasible for such complex systems. Leaf growth can be modelled via response curves to environmental conditions which are considered as 'meta-mechanisms' at a whole-plant degree of organisation. Response curves of leaf elongation rate to meristem temperature, water vapour pressure difference and soil water status were established in recombinant inbred lines (RILs) of maize in experiments carried out in the field or in the greenhouse. A QTL analysis was carried out on the slopes of these responses; most QTLs were specific for one response only. Each parameter of the ecophysiological model could then be computed as the sum of QTL effects, allowing calculation of parameters of new RILs, either virtual or existing. Leaf elongation rates of new RILs were simulated and were similar to measurements in a growth chamber experiment. This opens the way to the simulation of virtual genotypes, known only by their alleles, in any climatic scenario. Each genotype is therefore represented by a set of response parameters, valid in a large range of conditions and deduced from the alleles at QTLs. This approach, applied here to a relatively simple mechanism, might be extended to more complex plant modelling.
Virtual computer genotypes can be obtained from the modelling of plant responses to abiotic stresses. They might allow the behaviour of genotypes to be tested in silico.
Key words
Water deficit, tolerance, modelling, QTL, leaf growth, temperature.
Introduction
Leaf area is the main determinant of photosynthesis, via the well-documented relationship between light interception and biomass production (Monteith, 1977). Rapidly reaching a high proportion of absorbed light during spring is therefore an important trait for crop productivity. However, leaf area is also an important determinant of plant transpiration, so a common adaptive trait for drought tolerance is a reduced leaf growth under water deficit. Reduction in leaf expansion rate usually occurs before any reduction in photosynthesis (Boyer, 1970, Saab and Sharp 1989), or in the growths of other plant organs (Wesgate and Boyer 1985, Saab et al , 1990). It occurs in response to both air and soil water deficits (Ben Haj Salah and Tardieu, 1997, Serpe and Matthews 2000), resulting in a reduction in transpiration which contributes to avoid cell water stress even when the plant is in water deficit. On short timescales, reduction in leaf area has a similar role as stomatal closure that is to reduce the water flow through the plant, thereby flattening the gradient of water potential between the soil and the leaves. This avoids deleterious leaf water potentials in leaves. On longer timescales, a reduced leaf area saves soil water for later stages of plant development via the reduction in transpiration.
Breeding for tolerance to water deficit can therefore involve two opposite strategies: either select plants which reduce leaf area under water deficit in order to avoid plant water stress, or select plants with a maintained leaf area in order to maximise light interception under water deficit.
- Several genetic manipulations are based on the first strategy and increase plant survival under water deficit. For instance, Arabidopsis plants which overexpress a gene involved in the synthesis of abscisic acid (ABA) stay green longer and die after control plants (Iuchi et al., 2001). This is probably linked to the fact that an overproduction of ABA has a reducing effect on leaf area and stomatal conductance, and therefore on transpiration. The same sequence of events allow tomato plants which overexpress the transcriptional activator CBF1, DREB1B, to stay green longer than wild type plants (Hsieh et al., 2002). The second strategy consists of maintaining growth, photosynthesis and development during the whole cycle under dry conditions. Maize plants, obtained in a divergent selection for yield under dry conditions, have these characteristics and also maintain the number of kernels under water deficit (Bruce et al. 2002). This allows plants to accumulate a maximum biomass during dry periods, with the risk that the elevated transpiration associated with high photosynthesis causes total water depletion and plant death before the end of the crop cycle.
Which of these strategies best contributes to the tolerance of water deficit? The answer probably depends on the climatic scenario. The first, conservative, strategy is suited to most adverse scenarios in which saving water is essential. The second strategy is adapted to scenarios with shorter water deficits in which it is not crucial to save water, but to maximise growth at any stage of development even in moderately dry soil. Tolerance to water deficit therefore involves, at least in part, a balance between reduction in risk of severe stress, obtained by stomatal closure and reduction in leaf area, and conservation of the ability of the plant to accumulate biomass which involves leaf growth maintenance. Because rainfall varies year to year and periods of deficit can occur at any time of the crop cycle, more than seven years are usually necessary to explore the most likely climatic scenarios (Chapman et al. 2000). In addition, the genes that confer tolerance to stress at one stage of development differ from those conferring tolerance at another stage (Foolad 1999) so a given genotype should be tested in all likely climatic scenarios. The use of controlled environments reduces the climatic variability without reducing the number of scenarios to be tested. This suggests that an exclusively experimental procedure is not feasible. Simulation of the behaviour of genotypes in multiple climatic scenarios could be a useful tool to test the effects of gene transformation or allelic diversity on the tolerance to water deficit.
Can we model the genetic variability of leaf response to water deficit from known mechanisms and genes?
A conceptually simple strategy would consist of placing the effects of all involved genes in a regulatory network which simulates the effects of fluctuating environmental conditions on their transcript levels, on the amount and on the activities of corresponding proteins, and finally on the resulting phenotype. This strategy has been used for simulating relatively simple systems such as the behaviour of prokaryotes as a function of their environment (Gilman and Arkin, 2002), or the fate of Arabidopsis thaliana reproductive organs (Jack 2001). It has been considered as suitable to study the effects of water deficit on plants (Cushman and Bonhert, 2000). However, is our knowledge of the regulatory processes involved in the control of leaf growth precise enough for such a prediction? Several mechanisms are commonly considered to be involved in the control of leaf growth, and each of them is the object of a debate.
Changes in cell wall plasticity and in cell division rate.
Water deficit affects the plasticity of cell walls, thereby reducing leaf expansion rate (Matthews et al. 1984). The opposite occurs in roots, thereby allowing roots to partly maintain growth under water deficit in spite of a reduced turgor (Spollen and Sharp 1991, Wu et al 1996). There is now a consensus that cell wall stiffening is a major cause of the reduction in leaf growth (Cosgrove 1999, Tang and Boyer 2002). However, the molecular processes are not understood in a way comprehensive enough to allow modelling. Three gene families are the main molecular candidates for changes in cell wall properties. Expansins, which can induce cell wall extension in vitro, are believed to disrupt the hydrogen bonds between cell wall polymers (Cosgrove 1999). Their expression is appreciably affected by water deficit (Wu and Cosgrove 2001) but, because it is a large multigenic family, the correspondence between transcript levels and expansion rate is not straightforward (Caderas et al 2000). It is hypothesised that xyloglucan endotransglycosylase (XET) cuts and rejoins xyloglucan molecules which form tethers between cellulose microfibrils, but controversy remains on the exact role of this family of enzymes and even on their effects on growth (Cosgrove 1999). Finally, cell wall peroxidases act as a stiffening agent by catalysing cross links between phenolic groups in the cell wall. The cell-wall associated peroxidase activity is increased in Lolium leaves subjected to water deficit, but without clear demonstration of their role (Bacon et al. 1997). Other families of proteins may also be involved, such as endoglucanases (Yuan et al. 2001). Each of these families can involve several tens of genes whose individual effects are not known, and the interaction between families of genes is still less known.
Cell division rate in leaves of several species is clearly affected by water deficit in both dicot (Yegappan et al. 1882, Granier and Tardieu 1999) and monocot leaves (Schuppler et al. 1998, Tardieu et al. 2000). The limiting stage of the cell cycle probably precedes the entry in the S phase of the cycle, because cells of droughted leaves tend to accumulate in the G1 phase. In maize, the reduction in cell division rate due to several environmental conditions, especially temperature and water deficit, is clearly linked to the activity of a key enzyme of the cell cycle, the p34cdc2kinase (Granier et al. 2000). However, it is not linked to the amount of this enzyme, but probably to inhibitors of its activity (Schuppler et al. 1998). Again, the precise role of individual genes in the response to water deficit is insufficiently known to allow a predictive approach.
Cell turgor and osmotic adjustment.
Reduction in cell turgor has long been considered as a straightforward cause of the reduction in leaf growth under water deficit (Zhang et al., 1999). This is observed in very short-term experiments (Pritchard et al 1993), and follows the theoretical framework of Lockhart (1965). This interpretation of reductions in leaf expansion rate under water deficit would imply that osmotic adjustment in growing tissues is not complete enough to compensate the decrease in total water potential, so turgor would decrease in cells of droughted plants. The theory of a central role of turgor in the control of leaf growth has been discussed for the last thirty years (Green et al. 1971). Reductions in leaf elongation rate were observed in response to soil water deficit in spite of an unchanged turgor pressure (Matthews et al. 1984; Wesgate and Boyer 1985, Tang and Boyer 2002). The view that leaf growth under water deficit is unrelated to cell turgor would imply first that osmotic adjustment is rapid enough to compensate the decrease in total water potential in the leaf, and second that other mechanisms such as cell wall stiffening must be invoked to account for the reduced leaf expansion rate under water deficit. There is, therefore, a contradiction between two sets of papers which originate from different scientific communities and were obtained using different ways of calculating turgor and osmotic adjustment. Ideally, these two views could be challenged by direct measurements of cell turgor with a cell pressure probe, in growing zones of plants subjected to soil or air water deficits. However, few data are available, with contrasting results. Shackel (1987ab) and Serpe and Matthews (2000) observed a marked change in turgor of dicot leaves following a change in evaporative demand, with small changes in solute potential. In the same way, Spollen and Sharp (1991) observed a turgor decrease in maize roots subjected to a severe water stress (-1.5 MPa), contrasting with results of Pritchard et al (1991) in barley. If the importance of osmotic adjustment is still controversial, it is unlikely that one can predict in the short term the effect of genes involved in osmotic adjustment.
Hormone signalling
Abscisic acid (ABA) is widely believed to be a major contributor in the signalling pathway of water deficit and in the controls of plant transpiration and leaf growth. This view is reinforced by experiments in which the ABA biosynthesis pathway was affected (Iuchi et al. 2001, Borel et al. 2001) or in which artificial ABA was fed to plants (Zhang and Davies 1989, Ben Haj Salah and Tardieu 1997). Models involving ABA have been proposed for the controls of transpiration (Tardieu and Davies 1993, Dewar 2003) and of leaf expansion rate (Ben Haj Salah and Tardieu 1997). However, the picture is more complex when the effect of ABA is dissected genetically. The signalling pathways of ABA and ethylene overlap, because mutants affected in sensitivity to ABA are allelic with mutants of ethylene sensitivity (Beaudoin et al, 2000). A similar overlapping is observed between the signalling pathways of ABA and of sucrose (Leon and Sheen 2003). Furthermore, recent studies show that the respective roles of ABA and ethylene may have to be reconsidered in case of water deficit and that ABA might promote growth instead of inhibiting it in case of water deficit (Sharp 2002).
Can gene regulatory networks be used for predicting the response of leaf growth to water deficit?
In each of the above-mentioned mechanisms, widely believed to be involved in the response of leaf growth to water deficit, there still are fundamental debates on the effects of the considered functions or genes. Instead of simplifying the picture, the molecular dissection of each mechanism has tended to increase its complexity, to generate new debates and increase the number of involved genes because several families of up to 30 genes are involved in each single mechanism. Transcriptome analyses should help to analyse this complexity but provide no avenues for modelling plant behaviour. Another difficulty concerns the phenotypic analysis. When plant survival is analysed, the phenotype is qualitative with two possibilities (alive / dead), as in the gene regulatory network analysed by Gilman and Arkin (2002). When leaf growth is analysed, the phenotype is quantitative i.e. an infinite number of values of each variable and of combinations of variables are possible. The analysis of plant phenotype is necessarily based upon a theory, even implicit, which leads one to choose the variables to be measured, the time to measure them and the chosen levels of stress in experiments.
If models of behaviours of genotypes are to be developed, they will therefore be based on principles which differ from the gene regulatory networks, at least in the next years or decades.
"Meta-mechanisms " can be identified at plant or organ levels.
Crop modellers have long used another way of expressing the plant controls, that is to express a phenotypic trait at a given time (e.g. transpiration rate, expansion rate of organs or biomass accumulation) as a function of environmental inputs such as organ temperature, light irradiance or soil water potential. Relatively simple equations are used for this purpose, some of which are straightforward because they represent a physical process. For example, physical equations describing mass or heat fluxes, such as water transfer in the plant or energy balance of the leaf, have a known formalism. The parameters of equations have a physical value which characterises an environment (e.g. soil hydraulic properties) or a genotype (e.g. leaf albedo, hydraulic conductivity of the xylem). The difficulty in the use of these equations is to define the degree of simplification which is acceptable and to experimentally determine their parameters, but the formalism of equations is usually non-controversial.
Control equations have another status: they describe, for example, the response of growth to an environmental condition or the progression of development of the plant. These equations are based neither on physical concepts nor on the knowledge of gene action. They are therefore theoretically fragile and can be misleading. However, several examples suggest that control equations may have a value per se, such as that presented in Fig. 1 for the response of leaf elongation rate to meristem temperature. Although the combination of molecular mechanisms which leads to the response to temperature is not known, leaf elongation rate is linearly related to meristem temperature, and the same response curve applies to plants grown over several years in the field, in the greenhouse and in the growth chamber provided that the plant experiences no stress and no evaporative demand (Fig. 1a-c). The slope of this relationship is therefore a stable characteristic of the genotype, and differs between genotypes (Fig. 1d). In this example, it would be impossible to establish the gene regulatory network which controls the response of leaf elongation rate to temperature, but the quantitative analysis of the phenotype allows predicting the response of a genotype and comparing genotypes.
"Meta-mechanisms" are therefore control equations which can seem empirical at first sight. They are based on two sensible assumptions: (i) that the network of gene regulation is coordinated in such a way that a plant reacts in a predictable way to a given environmental condition, giving way to a mechanism at a higher level of organisation, and (ii) that a finite number of combinations of molecular mechanisms have been selected by evolution, so it is possible to analyse the responses to environmental conditions at the plant level. The legitimacy of these mechanisms at plant level rests on their stability over contrasting environmental scenarios, in addition to their compatibility with current physiological knowledge. Any relationship obtained during an experiment does not provide a control equation. In the example of Fig. 1, the relationship between night-time leaf elongation rate and air (instead of meristem) temperature is experimentally unstable. Neither is it stable during the day if the effect of evaporative demand is not taken into account (Ben Haj Salah and Tardieu 1996). Once a control equation (or a combination of control equations) is proposed, its validity is tested with conditions and genotypes different from those on which it was established (Reymond et al. 2003). The consistency of a putative model with kinematic analyses of growth and development analyses (Tardieu et al. 2000, Granier et al. 2000), of transcript levels (Pic et al. 2000) or of protein activity (Granier et al. 2000) can also be checked. Several potentially acceptable models are usually discarded during this process. Obtaining such models is time consuming but provides a generic characterisation of a series of genotypes in any environmental condition.
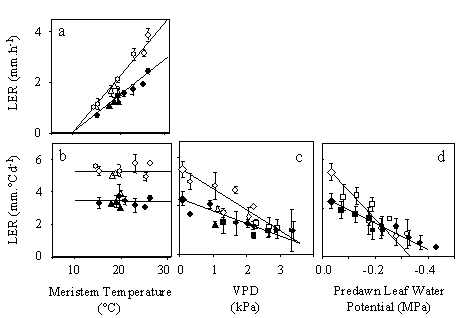
Figure 1. Dissection of the responses of leaf elongation rate (LER) to temperature, evaporative demand and soil water status in two typical RILs (open and filled symbols). a. LER per unit clock time, plotted against meristem temperature. b. LER per unit thermal time, plotted against meristem temperature. The mean LER is an estimate of parameter a of Eq. 1. c. LER per unit thermal time, plotted against meristem to air water vapor pressure difference (VPD) in well-watered plants. d. LER per unit thermal time during night periods, plotted against predawn leaf water potential.. Each symbol, one experiment in the field ( ‘ , ∇ ) or in the greenhouse ( Δ ¡ ‘ u). Redrawn from Reymond et al. (2003).
Bases for modelling leaf expansion rate and its response to environmental conditions
Expressing elongation rates per unit thermal time.
Thermal time is used in crop modelling to take into account the effect of temperature on plant development. We have proposed (Granier and Tardieu 1998) that its use can be extended to the calculation of rates with a sound mathematical basis. Leaf expansion rate and cell division rate respond linearly to organ temperature (Fig. 2), with an x-intercept which is most often common to several processes. For instance in sunflower, the x-intercepts for cell division rate, tissue expansion rate, duration of expansion, duration of cell division and leaf appearance rate have a common x-intercept of 5°C (Granier and Tardieu 1998). The same applies to maize (Ben Haj Salah and Tardieu 1995) and Arabidopsis thaliana (Granier et al. 2002). This leads to two equations. The first relates temperature to the rates of processes involved in leaf growth:
(1) dL / dt = a (T - T0)
where L is leaf length (or surface, or cell number), T is current temperature, a and T0 are the slope and the x-intercept of the relationship between dL/dt and T. The second relationship involves the reciprocal of the duration of the studied process:
(2) 1/d = b (T - T0)
where d is the time during which expansion (respectively cell division) occurs in a given leaf, or the time during which leaf initiation occurs on the apex. Because these equations account for relationships which have a broad value and apply to fluctuating as well as stable conditions (Fig. 2), they can be integrated and used for expression of leaf elongation rate per unit thermal time. At time d:
(3) ![]()
![]() is thermal time (unit : °Cd when calculated with a daily timestep). This implies that time, as sensed by plants, elapses more rapidly at high than at low temperature but that both durations and rates are independent of temperature if expressed in thermal time. This can be visualised in Fig. 1b in which maize leaf elongation rate becomes independent of meristem temperature if expressed per unit thermal time. This way of expressing rates is a powerful tool to analyse processes in naturally fluctuating conditions.
is thermal time (unit : °Cd when calculated with a daily timestep). This implies that time, as sensed by plants, elapses more rapidly at high than at low temperature but that both durations and rates are independent of temperature if expressed in thermal time. This can be visualised in Fig. 1b in which maize leaf elongation rate becomes independent of meristem temperature if expressed per unit thermal time. This way of expressing rates is a powerful tool to analyse processes in naturally fluctuating conditions.

Figure 2. Response to leaf temperature of leaf expansion rate (a), cell division rate (b) the reciprocal of the duration of expansion of the leaf (c) and the reciprocal of the duration of cell division (d). Data were collected in several experiments with sunflower plants in the field or in the greenhouse. Redrawn from Granier and Tardieu 1998.
High evaporative demand decreases leaf elongation rate of several species even in moist soil.
A clear effect of air water status has been observed on leaf expansion rate of maize (Ben Haj Salah and Tardieu 1996) and begonia (Serpe and Matthews 2000), in addition to the effect of soil water deficit. This effect cannot be observed in common growth chambers or greenhouses where water vapour pressure deficit (VPD) and light intensity are low. It is very clear in Mediterranean climates where VPD undergoes large variations (Ben Haj Salah and Tardieu 1996). A high and constant evaporative demand without soil water deficit caused approximately the same effect on the spatial distribution of RER as a change in soil water status between consecutive days. This was in spite of the fact that predawn leaf water potential and the concentration of ABA in the xylem sap were close to 0 (Ben Haj Salah and Tardieu 1997). This effect is species-dependent, for instance it was not observed in sunflower. It is also genotype-dependent in maize (Reymond et al. 2003).
Response to soil water deficit
Leaf expansion rate decreases with soil water potential in nearly all species. In dicot plants such as sunflower or pea, the effect of soil water deficit is greatest during the early stages of leaf development, during which both relative expansion rate and cell division rate are maximum (Lecoeur et al. 1994, Granier et al. 1999). In monocot plants such as maize, this effect seems largely independent of the stage of leaf development as leaf elongation rate is affected in a reproducible way in several experiments at different stages. The spatial distribution is affected, also in a reproducible way, at all distances from the leaf insertion point, resulting in a reduction of the elongating zone of the leaf. Water deficit affects both tissue expansion and cell division, however expansion is slightly more affected than cell division, resulting in smaller cells in the mature zone of the leaf. A large genetic variability of the sensitivity of leaf expansion rate to soil water deficit was observed in maize (Reymond et al. 2003).
Analysis of the genetic variability of the responses of leaf elongation rate to water deficit.
The final leaf length changes with environmental conditions, but the response of leaf elongation rate to temperature, evaporative demand and soil water status are stable for a given genotype and apply to field as well as to controlled conditions (Figs. 1, 2). They can therefore be considered as intrinsic characteristics of the considered genotype, and be analysed in a genetic analysis.
We have proposed recently that a QTL analysis can be carried out on parameters of the response curves of maize leaf elongation rate to environmental conditions, thereby dealing with an adaptive trait using an explicit treatment of the genotype x environment interaction (Reymond et al. 2003). QTLs were determined for sensitivities of leaf elongation rate to meristem temperature, leaf to air vapour pressure deficit and soil water status. It was shown earlier that light intensity has no direct effect on maize leaf elongation rate at this timescale, and essentially acts via its effect on leaf-to-air VPD (Ben Haj Salah and Tardieu 1996). Each slope of the response curves presented in Fig. 1 was established in a series of experiments:
- The response of leaf elongation rate to meristem temperature was considered during night periods in experiments without soil water deficit, when the evaporative demand was null and the leaf elongation rate only depended on meristem temperature.
- The response to evaporative demand was considered during day periods without soil water deficit, while meristem temperature, light intensity and evaporative demand fluctuated with time. The sensitivity to evaporative demand was then estimated via the slope of the response curve of leaf elongation rate to meristem-to-air VPD.
- The response to soil water deficit was analysed during night periods. The slope of the response curve of leaf elongation rate to predawn leaf water potential was an estimate of the sensitivity to soil water status.
The three responses can be combined in a model with three parameters (Tardieu et al. 2000):
(4) dL/dt = (T-T0) (a + b VPD -c Ψ)
where dL/dt is leaf elongation rate, T is meristem temperature; a and To are the slope and the x-intercept of the relationship between meristem temperature and leaf elongation rate; b (negative value) is the slope of the relationships between leaf elongation rate (in mm °Cd-1) and VPD; c (negative value) is the slope of the relationship between leaf elongation rate (in mm °Cd-1) and soil water potential.
Each parameter of the response curves was analysed in a QTL analysis. The QTL models accounted for 55, 47 and 43 % of the genetic variabilities of parameters a, b and c, respectively. QTLs of each parameter had few colocations, suggesting that they were controlled independently by the genome. Each parameter of Eq. 4 was then expressed as a sum of QTL effects. The resulting model, which combined QTL analysis and ecophysiological modelling, allowed predicting the behaviour of new recombinant inbred lines (RILs) under a new climatic scenario, although these new RILs were only known by their alleles at QTL positions. This combination of models was tested on 13 lines not involved in the construction of the QTL models. Leaf elongation rates measured in a growth chamber experiment were compared with those predicted by the model, using measured temperature, VPD and soil water potential as inputs (Fig. 3). Plants were subjected to a near-zero evaporative demand during the night, and to varying evaporative demands at a constant meristem temperature during the day (Fig. 3a). Examples of predicted and measured time courses of leaf elongation rates of 5 RILs are presented in Fig. 3bc. Leaf elongation rate had similar time courses in modeled and observed data. It decreased in three steps during the night, simultaneously with temperature (periods 1, 2, 3). It decreased steeply when lights were turned on and temperature was returned to 28°C. It partly recovered and stabilized under the low VPD (period 4). It decreased afterwards in two steps simultaneously with VPD (periods 5 and 6). The model predicted differences in elongation rates observed between RILs during the night at all temperatures. It also predicted the difference in response to evaporative demand during the day : one RIL (thin line) had a low response compared with the others, consistent with predicted values.
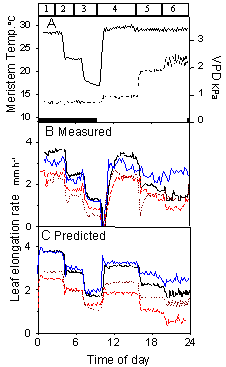
Figure 3. Time courses of measured and modeled leaf elongation rates during a climatic scenario in the growth chamber. Four recombinant inbred lines (RILs) are presented, which were not taken into account in the detection of QTLs. The modeled values were obtained from the ecophysiological model (Eq. 4) whose parameters were calculated as a sum of QTL effects with the genetic models. A. Change with time of meristem temperature (plain line) and VPD (dotted line). Numbers on the top of the panel represent periods, identified for better legibility in the text. Black bars on the bottom of the panel indicate the night periods. B, Leaf elongation rate measured with LVDTs, averaged on 2 or more plants of each RIL. Each line style represents a RIL. C, modeled leaf elongation rate for the same RILs. ). Redrawn from Reymond et al. (2003).
Conclusion
How can one link the physiological and genetic analyses of the control of leaf growth under changing environmental conditions? The view presented here suggests that aggregating all the available knowledge about the actions of genes into a model is not feasible for the time being, and that this may well be the case for a long time. We rather propose that the phenotype of a given genetic line might be 'footprinted' via a vector of parameters of models, and that a genetic analysis of parameters can be a useful avenue for modelling the genotype x environment interaction.
It must be recognised that the exercise presented here for coupling genetic and ecophysiological analyses applies to a relatively simple case. First it is carried out on a single phenotypic character, leaf elongation rate, which has to be combined with many other characters in order to predict the plant architecture, transpiration and biomass production. However, the recent work of Chapman et al (2003) suggests that such an integration of mechanisms is possible and might allow one to evaluate plant breeding strategies with crop models. Second, the 'virtual genotype' exercise was carried out in a single mapping population of recombinant inbred lines, while one would expect the method to apply to a wider genetic range. However, we have suggested elsewhere (Tardieu 2003) that the method can also apply to transgenic plants. We are presently applying it to other mapping populations (W. Sadok, C. Welcker and F. Tardieu unpublished data). It is possible to imagine that studies of association genetics, carried out on large panels of genetically unrelated lines could allow one to reconstruct the phenotype of plants in climatic scenarios for a large range of genetic variability.
References
Bacon MA, Thompson DS, Davies WJ (1997) Can cell wall peroxidase activity explain the leaf growth response of Lolium temulentum L. during drought ? Journal of Experimental Botany 48 2075-2085.
Boyer JS (1970) Leaf enlargement and metabolic rates in corn, bean and sunflower at various leaf water potential. Plant Physiology 46, 233-235
Beaudoin N, Serizet C, Gosti F, Giraudat J (2000) Interactions between abscisic acid and ethylene signaling cascades. Plant Cell 12, 1103-1115.
Ben Haj Salah H, Tardieu F (1997) Control of leaf expansion rate of droughted maize plants under fluctuating evaporative demand. A superposition of hydraulic and chemical messages? Plant Physiology 114, 893-900.
Ben Haj Salah H, Tardieu F (1996) Quantitative analysis of the combined effects of temperature, evaporative demand and light on leaf elongation rate in well-watered field and laboratory-grown maize plants. Journal of Experimental Botany 304, 1689-1698.
Borel C, Frey A, Marion-Poll A, Tardieu F, Simonneau T (2001) Does engineering ABA synthesis in N. plumbaginifolia modify stomatal response to drought ? Plant, Cell and Environment, 24, 477-489.
Bruce WB, Edmeades GO, Barker TC (2002) Molecular and physiological approaches to maize improvement for drought tolerance. Journal of Experimental Botany 53, 13-25.
Caderas D. Muster M, Vogler H, Mandel T, Rose JCK, Mc Queen Mason S, Kuhlemeier C (2000) Limited correlatioon between expansin gene expression and elongation growth rate. Plant Physiology 123, 1399-1413.
Chapman SC, Cooper M, Hammer GL, Butler DG (2000) Genotype by environment interactions affecting grain sorghum. II Frequencies of different seasonal patterns of drought stresses are related to location effects on hybrid yields Australian Journal of Agricutural Research 51, 209-221
Chapman SC, Cooper M, Podlich D, Hammer GL (2003) Evaluating plant breeding strategies by simulating gene action and dryland environment effects. Agronomy Journal 95, 99-113.
Cosgrove DJ (1999) Enzymes and other agents that enhance cell wall extensibility. Annual Review of Plant Physiology and Plant Molecular Biology 50, 391-417.
Cushman J.C., Bonhert H.J. (2000) Genomic approaches to plant stress tolerance. Current Opinion in Plant Biology 3, 117-124.
Dewar RC (2002) The Ball-Berry-Leuning and Tardieu-Davies stomatal models : synthesis and extension within a spatially aggregated picture of guard cell function. Plant Cell Environment 25, 1383-1398
Foolad MR (1999) Comparison of salt tolerance during seed germination and vegetative growth in tomato by QT mapping. Genome 42, 727-734
Gilman A, Arkin AP (2002) Genetic “code” : representations and dynamical models of genetic components and networks. Annual Review of Genomics and Human Genetics 3, 341-369.
Granier C, Tardieu F (1998). Is thermal time adequate for expressing the effects of temperature on sunflower leaf development ? Plant Cell Environment 21, 695-703.
Granier C Tardieu F (1999) Water deficit and spatial pattern of leaf development. Variability in responses can be simulated using a simple model of leaf development. Plant Physiology 119, 609-620.
Granier Ch, Inzé D, Tardieu F. (2000) Spatial distribution cell division rate can be deduced from that of P34cdc2 kinase activity in maize leaves grown in contrasting conditions of temperature and water status. Plant Physiology, 124, 1393-1402.
Green PB, Erickson R, Buggy J (1971) Metabolic and physical control of cell elongation rate. In vivo studies in Nitella. Plant Physiology 47, 423-430.
Hsieh TH, Lee JT, Charng YY, Chan MT ,2002. Tomato Plants Ectopically Expressing Arabidopsis CBF1 Show Enhanced Resistance to Water Deficit Stress. Plant Physiology, 130, 618-626.
Iuchi S, Kobayashi M, Taji T, Naramoto M, Seki M, Kato T, Tabata S, Kakubari Y, Yamaguchi-Shinozaki K, Shinozaki K. (2001) Regulation of drought tolerance by gene manipulation of 9-cis-epoxycarotenoid dioxygenase, a key enzyme in abscisic acid biosynthesis in Arabidopsis. Plant Journal 27, 325-333.
Jack T (2001) Relearning our ABCs : new twits on an old model. Trends Plant Sci. 6, 3110-3116
Lecoeur J, Wery J, Turc O, Tardieu F (1995) Expansion of pea leaves subjected to short water deficit : cell number and cell size are sensitive to deficit at different periods of leaf development. Journal of Experimental Botany 46, 1093-1101
Leon P, Sheen J (2003) Sugar and hormone connections. Trends in Plant Science 8, 110-116.
Matthews MA, Van Volkenburgh E, Boyer JS (1984) Acclimation of leaf growth to low water potentials in sunflower. Plant, Cell and Environment 7, 199-206.
Monteith J. L. (1977) Climate and the efficiency of crop production in Britain. Philosophical Transactions of the Royal society, London B 281, 277-294
Pic E, Teyssendier de la Serve B, Tardieu F, Turc O (2002) Leaf senescence induced by mild water deficit follows the same sequence of macroscopic, biochemical and molecular events as monocarpic senescence in pea (Pisum sativum L.) Plant Physiology 28, 236-246.
Pritchard, J., R.G. Win Jones and A.D. Tomos. 1991. Turgor, growth and rheological gradients of cereal roots and the effect of osmotic stress. Journal of Experimental Botany 42, 1043-1049.
Reymond M., Muller B., Leonardi A., Charcosset A., Tardieu F (2003) Combining quantitative trait loci analysis and an ecophysiological model to analyse the genetic variability of the responses of leaf growth to temperature and water deficit. Plant Physiology. 131, 664-675.
Saab IN, Sharp RE (1989) Non-hydraulic signals from maize roots in drying soil : inhibition of leaf elongation but not stomatal conductance. Planta: 466-474.
Saab IN, Sharp RE, Pritchard J (1990) Increased endogenous abscisic acid maintains primary root growth and inhibits shoot growth of maize seedlings at low water potentials. Plant Physiology 93: 1329-1336 .
Schuppler U, He PH, John PCL, Munns R (1998) Effect of water stress on cell division and cell-division-cycle 2-like cell cycle kinase activity in wheat leaves. Plant Physiology 117, 667-678.
Shackel KA, Matthews MA, Morrison JC (1987). Dynamic relation between expansion and cellular turgor in growing grape (Vitis vinifera L.) leaves. Plant Physiology 84, 1166-1171.
Shackel KA (1987) Direct measurements of turgor and osmotic potential in individual epidermal cells. Plant Physiology 83, 719-722.
Sharp RE (2002) Interaction with ethylene : changing views on the role of abscisic acid in root and shoot growth responses to water stress. Plant Cell Environment 25, 211-222/
Serpe MD, Matthews MA (2000) Turgor and cell wall yielding in dicot leaf growth in response to changes in relative humidity. Australian Journal of Plant Physiology 27, 1131-1140.
Spollen WG, Sharp RE (1991) Spatial distribution of turgor and root growth at low water potentials. Plant Physiology 96, 438-443
Tang AC, Boyer JS (2002) Growth-induced water potentials and the growth of maize leaves. Journal of Experimental Botany 368, 489-503.
Tardieu F (2003) Virtual Plants : modelling as a tool for the genomics of tolerance to water deficit. Trends in Plant Science 8, 9-14.
Tardieu F, Davies WJ (1993) Integration of hydraulic and chemical signalling in the control of stomatal conductance and water status of droughted plants. Plant, Cell and Environment, 16, 341-349.
Tardieu F, Reymond M, Hamard Ph, Granier Ch Muller B (2000) Spatial distributions of expansion rate, cell division rate and cell size in maize leaves. A synthesis of the effects of soil water status, evaporative demand and temperature. Journal of Experimental Botany 51, 1505-1514.
Westgate ME, Boyer JS (1985) Osmotic adjustment and the inhibition of leaf, root, stem and silk growth at low water potentials in maize. Planta 164, 540-549.
Wu, Y. Cosgrove, D.J. (2000) Adaptation of roots to low water potentials by changes in cell wall extensibility and cell wall proteins. Journal of Experimental Botany 51, 1543-1553.
Yegappan TM, Paton DM, Gates CT, Muller WJ (1982) Water stress in sunflower. II Effects on leaf cells and leaf area. Annals of Botany 49, 63 68.
Yuan S, Wu Y, Cosgrove DJ (2001) A fungal endoglucanase with plant cell wall extension activity. Plant Physiology 127, 324-333.
Zhang J, Nguyen HT, Blum A (1999) . Genetic analysis of osmotic adjustment in crop plants. Journal of Experimental Botany 50, 291-302.
Zhang J, Davies WJ (1990) Does ABA in the xylem sap control leaf growth in soil-dried maize and sunflower plants? Journal of Experimental Botany, 41, 1125-32.

Search Site
- Crop Science
- Help/Feedback
- Contact us
- Sitemap