

Identification of genomic regions associated with durable stripe rust resistance in wheat line 11IBWSN50
1 The University of Sydney, Plant Breeding Institute, PMB11, Camden NSW 2570, Australia.
Email krsna@camden.usyd.edu.au
2 CIMMYT Int., Apdo. Postal 6-641,06600, Mexico DF, Mexico.
3 BiometricsSA, University of Adelaide, Private Mail Bag 1, Glen Osmond, SA, 5064.
Stripe rust, caused by P. striiformis f. sp. tritici is an important disease of wheat that causes wide spread losses in many countries. This study was aimed to identify molecular markers linked to adult plant resistance to stripe rust in wheat (Triticum aestivum L), which could ease the incorporation of such resistance into newer wheat germplasm. A single-backcross derived F6 population from the cross Avocet S x 11IBWSN50 was used. Simple single marker regression analysis identified 2 genomic regions, 7DS and 4BL, involved in reduction of stripe rust severity. These findings putatively establish Yr18/Lr34 association on 7DS in 11IBWSN50. This also indicated that stripe rust resistance controlled by a genomic region on chromosome 4BL may be associated with leaf rust resistance similar to that of Pavon 76.
Wheat genotype 11IBWSN50 has shown durable resistance to both stripe rust and leaf rust diseases and hence is a valuable source for breeding rust resistant cultivars. This study aims at elucidating genetic and molecular basis of rust resistance in 11IBWSN50. Preliminary results suggested the involvement of chromosome arms 4BL and 7DS in reducing stripe rust severity. Two other genomic regions on chromosome arms 2AS and 1BL showed genetic associations with low stripe rust response during bulked segregant analyses, however, the associations did not turn out be statistically significant in QTL analysis.
Key words
QTL mapping-adult plant resistance-microsatellite marker-regression analysis
Introduction
Stripe rust of wheat, caused by Puccinia striiformis f. sp. tritici, causes severe damage to wheat production throughout the world periodically. Deployment of durable sources of resistance, and/or pyramided resistance genes, may slow down evolutionary process in pathogen populations (Jacobs and Parlevliet 1993). Slow rusting type of adult- plant resistance (APR) is well documented in several host/pathogen systems (Kaur et al. 2000, Jensen et al. 1999, Singh and Rajaram 1994) and has been investigated as a durable source of rust resistance in wheat (Bariana and McIntosh 1995). The inheritance of APR is often polygenic and additive, as indicated by continuous phenotypic variation.
Characters exhibiting continuous variation are called quantitative traits. The two reasons for this continuous variation are simultaneous segregation of many genes controlling the trait, and/or environmental influences on the expression of the trait (Falconer and Mackay 1996). Many important agronomic traits are quantitative and the loci affecting them are commonly referred to as quantitative trait loci (QTL). Biometrical methods have been traditionally used to study quantitative traits. However, the genetic basis of quantitative traits remained unclear until the advent of genetic maps based on phenotypically neutral DNA markers. Microsatellites or simple sequence repeats (SSRs) are a useful marker system with the advantage of being efficient, stable, co dominant, abundant and distributed throughout the genome. A SSR map of wheat with 279 marker loci was constructed by RÖder et al. (1998). Since then, genetic associations of many SSR markers with stripe rust resistance genes have been reported (Bariana et al. 2002).
11IBWSN50 (Pedigree Robin/ Chanate/ Bluebird/ Norteno 67) showed high levels of APR to both stripe rust and leaf rust diseases of wheat. Genetic analysis of a single backcross-derived (BC1) population from a cross involving 11IBWSN50 and stripe rust susceptible parent Avocet S indicated that 3 to 4 genes are involved in conditioning low stripe rust response. The objective of this study was to identify genomic regions conditioning low stripe rust response.
Methods
Population and phenotyping
The BC1-derived (Avocet*2/11IBWSN50) F6 population was tested under field conditions in Australia and Mexico. Puccinia striifomis f.sp. tritici (Pst) pathotypes used at both sites were virulent on seedling resistance genes Yr6 and YrA possessed by 11IBWSN50. Stripe rust severity was measured as percentage of leaf area infected in both Australia (Plant Breeding Institute-Cobbitty) and CIMMYT, Mexico.
DNA extraction and PCR conditions
DNA extraction from each line plus the parents was done using CTAB method (Doyle et al. 1990). PCR amplifications were done in 20μL volumes containing 50ng genomic DNA, 0.2mM dNTPs, 0.5μM of each primer, 1.5mM MgCl2, 1x PCR buffer and 1 U Taq DNA polymerase. A standard Touch Down PCR profile (Don et al. 1991) was used for all PCR amplifications. PCR products were mixed with 20μL of sequencing dye (98% formamide, 10mM EDTA pH 8 and 0.025% bromophenol blue-xylene cyanole).
Gel electrophoresis and gel staining
The PCR products were denatured at 95°C for 3 minutes and placed on a cold block until use. Each sample (4μL) was loaded on to pre-heated sequencing gel. Electrophoresis was performed at 60W for 90 minutes using a BIO-RAD Power Pack 3000 in 1X TBE buffer. Gels were silver-stained according to the protocol of Bassam et al. (1991).
Marker development
A total of 400 microsatellite primers from Röder et al. (1998) and the Wheat Microsatellite Consortium (WMC) were screened on parents 11IBWSN50 and Avocet S to identify parental polymorphisms. A leaf rust susceptible line WA1382 was included in the parental screening as well. Bulk segregant analysis (Michelmore et al. 1991) was performed on bulked DNA from 10 highly resistant and 8 highly susceptible individuals. The polymorphic and low stripe rust response linked markers were mapped on the entire population. The concentration of each individual component of the bulk was 30ng/μL.
Statistical analysis
Simple single marker regression analysis was performed to determine phenotype:genotype correlations.
Results
The backcross derived population showed continuous variation for stripe rust response in field experiments at both PBIC and CIMMYT. The distribution of stripe rust severity at PBIC during 2002 is shown in Figure 1.
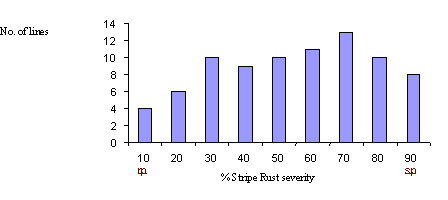
Figure 1. Distribution of 82 BC1F6 lines derived from Avocet S*2/ 11IBWSN50 PBIC during 2002.
Of 400 markers used, 38 showed polymorphism between Avocet S and 11IBWSN50. Eighteen of which covered 4 genomic regions (1B, 2A, 4B and 7D) and showed association with the DNA bulks based on stripe rust response. These polymorphic and linked markers were then screened on the entire population. Single marker regression analysis indicated a QTL by location interaction, with the CIMMYT trial detecting QTL linked to Xgwm495 on chromosome 4BL (LOD score 1.91), while the Australian trials in both years detected QTLs linked to Xgwm130 (LOD 1.36) and Xgwm295 (LOD 1.52) on chromosome 7DS. These markers (Xgwm495, Xgwm295 (Fig. 2) and Xgwm130) also showed polymorphism between 11IBWSN50 and a leaf rust susceptible parent WA1382 (Fig. 2).

Figure 2 Bulked segregant analysis results with Xgwm495 (left) and Xgwm295 (right) with leaf rust DNA bulks. Lane 1 susceptible parent, lane 2 resistant parent, lane 3 resistant bulk, gap and lane 4 susceptible bulk (Arrows indicate associated bands).
Conclusion
Regression analysis indicated the involvement of genomic regions on chromosome 4BL and 7DS in reducing stripe rust severity. The LOD scores were below the normally accepted threshold of 3, probably because of the modest sample size and higher levels of heterozygoisty in the population. Tests on a larger population may provide better resolution. The regions identified are, however, consistent with other recent studies. William et al. (2001) showed the presence of QTLs for stripe rust resistance on chromosome 4BL (Xgwm495) in Pavon 76 and 7DS (Xgwm130) in Parula. Linkage mapping on Parula indicated an association of Xgwm130 with leaf tip necrosis implicating the presence of Lr34/Yr18. Further, the same study associated a decrease in leaf rust severity with the Xgwm495-4B in Pavon 76. In addition, a recent report by Suenaga et al. (2003) indicated that Xgwm295 is also associated with Lr34/Yr18. Based on the above evidence, it is suggested that the QTL present on 7DS in 11IBWSN50 is linked to Lr34/Yr18. The present results obtained from bulk segregant analysis on the leaf rust resistance DNA bulks also indicated this association, at least putatively for time being. Lr34/Yr18 linked leaf tip necrosis was also observed in 11IBWSN50 (H.S. Bariana unpublished results). The 7DS and 4BL leaf rust/ stripe rust associations reported here are putative because the polymorphic and linked markers Xgwm130-7DS, Xgwm295-7DS and Xgwm495-4B have to be screened across the entire population to make definite conclusions.
Two genomic regions (1BL and 2AS) identified being associated with stripe rust response variation during bulked segregant analyses were not statistically significant when single marker regression analysis was performed. This is probably due to limitation of single marker analysis often referred as “single marker syndrome”. Because of the limited number of markers on these chromosomes and higher levels of heterozygosity in population, other analysis systems like interval mapping were not appropriate. Singh et al. (1998) reported the presence of Yr29/Lr46 in chromosome 1BL of Pavon 76. 11IBWSN50 was distributed in the 11th International Bread Wheat Screening Nursery together-with Pavon 76. 11IBWSN50 and Pavon 76 both possess seedling rust resistance genes Yr6, Lr13 and Sr30 (H.S. Bariana unpublished results). It is likely that these genotypes share pedigree and therefore 11IBWSN50 may carry Yr29.
References
Bassam BJ, Caeteno-Anolles G and Gresshof PM (1991). Fast and sensitive silver staining of DNA in Polyacrylamide gels. Anal. Biochem.196, 80-83.
Bariana HS, Brown GN, Ahmed NU, Khatkar S, Conner RL, Wellings CR, Haley S, Sharp PJ and Laroche A (2002). Characterization of Triticum vavilovii-derived stripe rust resistance using genetic, cytogenetic and molecular analyses and its marker assisted selection. Theor. Appl. Genet. 104, 315-320.
Bariana HS and McIntosh RA (1995). Genetics of adult plant stripe rust resistance in four Australian wheats and the French cultivar Hybride-de-Bersee. Plant Breeding 114, 485-491.
Doyle JJ and Doyle JL (1990). Isolation of plant DNA from fresh tissue. Focus 12, 13-15.
Don RH. (1991). "Touch down" PCR to circumvent spurious priming during gene amplification. Nucleic Acids Res 19, 4008.
Falconer DS and Mackay TFC (1996). Introduction to Quantitative Genetics. Longman Group Ltd, Edinburgh, 464 pp.Jacobs TH and Parlevliet JE (1993). Durability of Disease Resistance. Kluwer Academic Publication., Dordrecht. Genetics 149, 2007-2023.
Jensen BD, Hockenhull J and Munk L (1999). Seedling and adult plant resistance to downy mildew (Peronospora parasitica) in cauliflower (Brassica oleracea var. botrytis). Plant Pathology 48, 604-612.
Kaur Mandeep, Saini RG, Kanwal Preet (2000). Adult plant leaf rust resistance from 111 wheat (Triticum aestivum L.) cultivars. Euphytica 113, 235-243.
Michelmore RW, Paran I and Kesseli RV (1991). Identification of markers linked to disease resistance genes by bulked segregant analysis: a rapid method to detect markers in specific genomic regions by using segregating populations. Proc. Natl. Acad. Sci.USA 88, 9828-9832.
Röder MS, Korzun V, Tixier MH, Leroy P and Ganal MW (1998). A microsatellite map of wheat. Genetics 149, 2007-2023.
Saini RG, Kaur M, Singh B, Sharma S, Nanda GS, Nayar SK, Gupta AK and Nagarajan K (2002). Lr48 and Lr49, novel hypersensitive adult plant leaf rust resistance genes in wheat (Triticum aestivum L.). Euphytica 124, 365-370.
Singh, R. P., A. Mujeeb-Kazi, and J. Huerta-Espino. 1998. Lr46: A gene conferring slow-rusting resistance to leaf rust in wheat. Phytopathology 88(9):890-894.
Singh RP and Rajaram S (1994). Genetics of adult plant resistance to stripe rust in ten spring bread wheats. Euphytica 72, 1-7.
Suenaga K, Singh RP, Huerta-Espino J and William HM (2003). Microsatellite markers for genes Lr34/Yr18 and other quantitative trait loci for leaf rust and stripe rust resistance in Bread Wheat. Phytopathology 93, 881-890.
William M, Singh RP, Huerta J, Palacios G, Suenaga K and D. Hoisington (2001). Charecterization of durable rust resistance genes with molecular markers in wheat. Proceedings of CIMMYT-Kronstad symposium.

Search Site
- Crop Science
- Help/Feedback
- Contact us
- Sitemap